

The C-terminal domain of the Arabinosyltransferase Mycobacterium tuberculosis EmbC is a lectin-like carbohydrate binding module
- PMID: 21383969
- PMCID: PMC3044687
- DOI: 10.1371/journal.ppat.1001299
The C-terminal domain of the Arabinosyltransferase Mycobacterium tuberculosis EmbC is a lectin-like carbohydrate binding module
Abstract
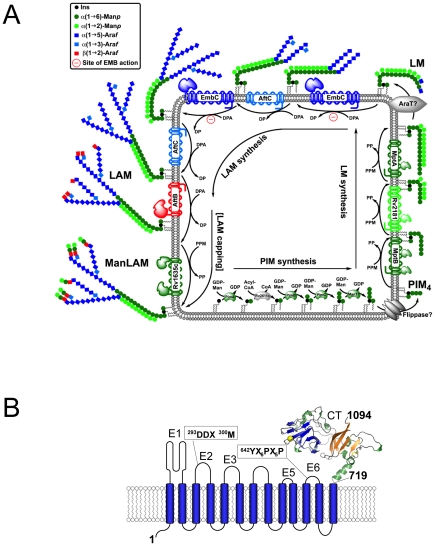
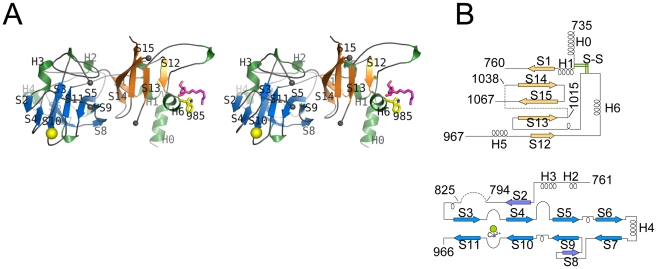
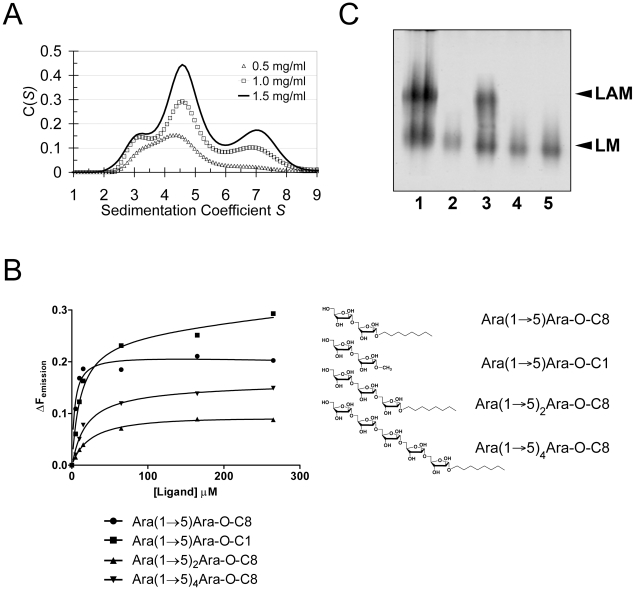
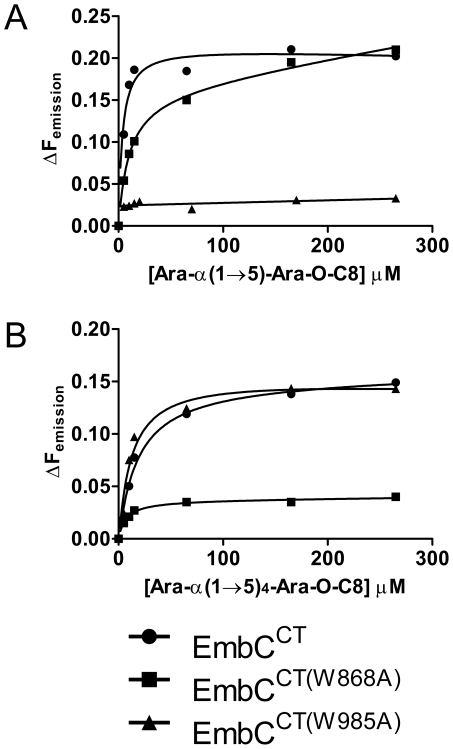
The D-arabinan-containing polymers arabinogalactan (AG) and lipoarabinomannan (LAM) are essential components of the unique cell envelope of the pathogen Mycobacterium tuberculosis. Biosynthesis of AG and LAM involves a series of membrane-embedded arabinofuranosyl (Araf) transferases whose structures are largely uncharacterised, despite the fact that several of them are pharmacological targets of ethambutol, a frontline drug in tuberculosis therapy. Herein, we present the crystal structure of the C-terminal hydrophilic domain of the ethambutol-sensitive Araf transferase M. tuberculosis EmbC, which is essential for LAM synthesis. The structure of the C-terminal domain of EmbC (EmbC(CT)) encompasses two sub-domains of different folds, of which subdomain II shows distinct similarity to lectin-like carbohydrate-binding modules (CBM). Co-crystallisation with a cell wall-derived di-arabinoside acceptor analogue and structural comparison with ligand-bound CBMs suggest that EmbC(CT) contains two separate carbohydrate binding sites, associated with subdomains I and II, respectively. Single-residue substitution of conserved tryptophan residues (Trp868, Trp985) at these respective sites inhibited EmbC-catalysed extension of LAM. The same substitutions differentially abrogated binding of di- and penta-arabinofuranoside acceptor analogues to EmbC(CT), linking the loss of activity to compromised acceptor substrate binding, indicating the presence of two separate carbohydrate binding sites, and demonstrating that subdomain II indeed functions as a carbohydrate-binding module. This work provides the first step towards unravelling the structure and function of a GT-C-type glycosyltransferase that is essential in M. tuberculosis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- World Health Organisation. 2009. Global Tuberculosis Control: a short update to the 2009 report ( http://www.who.int/entity/tb/publications/2009/factsheet_tb_2009update_d...)
-
- Harries AD, Dye C. Tuberculosis. Ann Trop Med Parasitol. 2006;100:415–431. - PubMed
-
- Jain A, Mondal R. Extensively drug-resistant tuberculosis: current challenges and threats. FEMS Immunol Med Microbiol. 2008;53:145–150. - PubMed
-
- Crick DC, Mahapatra S, Brennan PJ. Biosynthesis of the arabinogalactan-peptidoglycan complex of Mycobacterium tuberculosis. Glycobiology. 2001;11:107R–118R. - PubMed
-
- Briken V, Porcelli SA, Besra GS, Kremer L. Mycobacterial lipoarabinomannan and related lipoglycans: from biogenesis to modulation of the immune response. Mol Microbiol. 2004;53:391–403. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
