

The bZIP transcription factor MoAP1 mediates the oxidative stress response and is critical for pathogenicity of the rice blast fungus Magnaporthe oryzae
- PMID: 21383978
- PMCID: PMC3044703
- DOI: 10.1371/journal.ppat.1001302
The bZIP transcription factor MoAP1 mediates the oxidative stress response and is critical for pathogenicity of the rice blast fungus Magnaporthe oryzae
Erratum in
-
Correction: The bZIP Transcription Factor MoAP1 Mediates the Oxidative Stress Response and is Critical for Pathogenicity of the Rice Blast Fungus Magnaporthe oryzae.PLoS Pathog. 2019 Nov 20;15(11):e1008196. doi: 10.1371/journal.ppat.1008196. eCollection 2019 Nov. PLoS Pathog. 2019. PMID: 31747447 Free PMC article.
Abstract
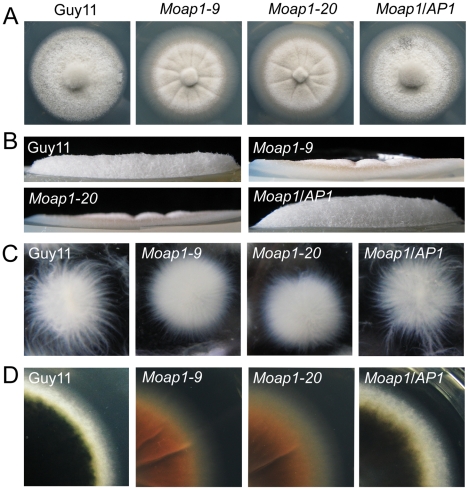
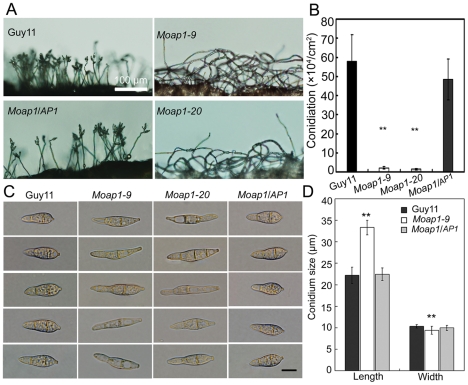
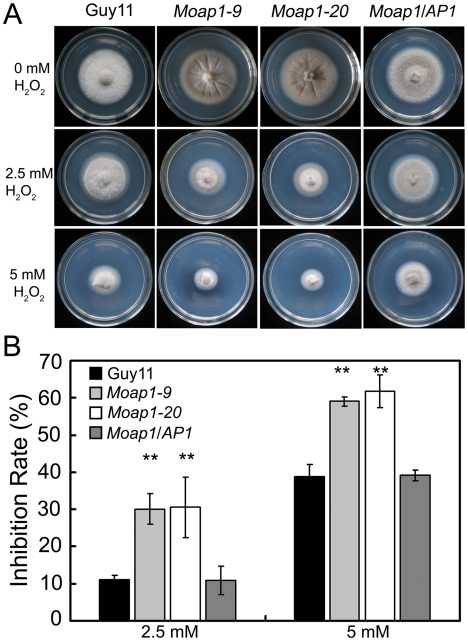
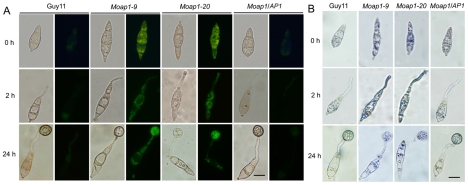
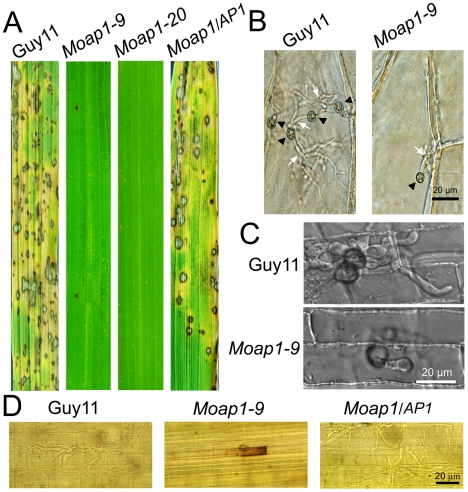
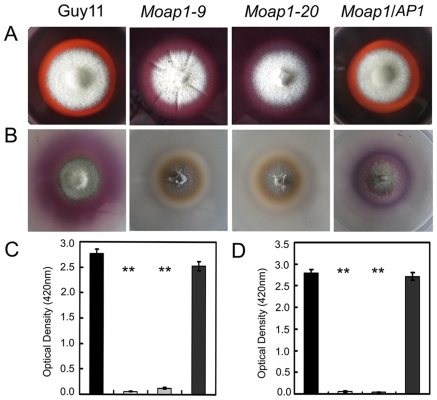
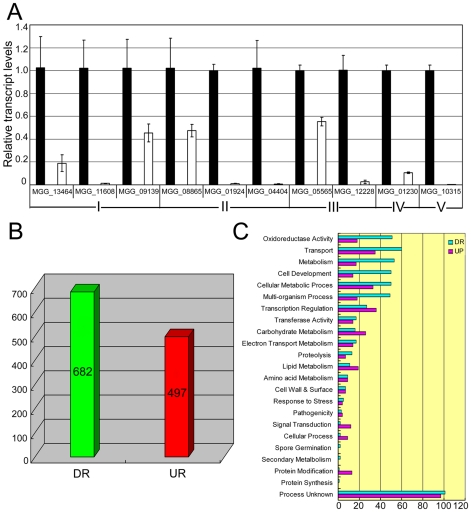
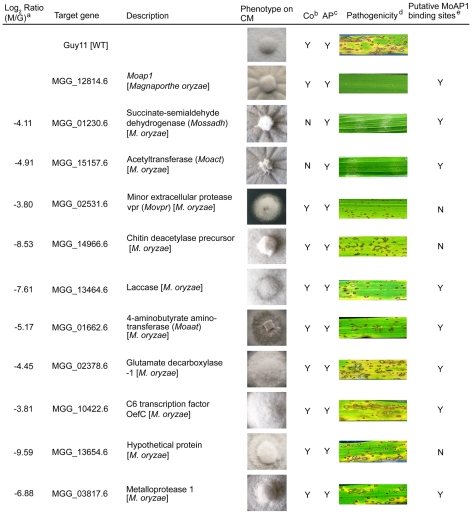
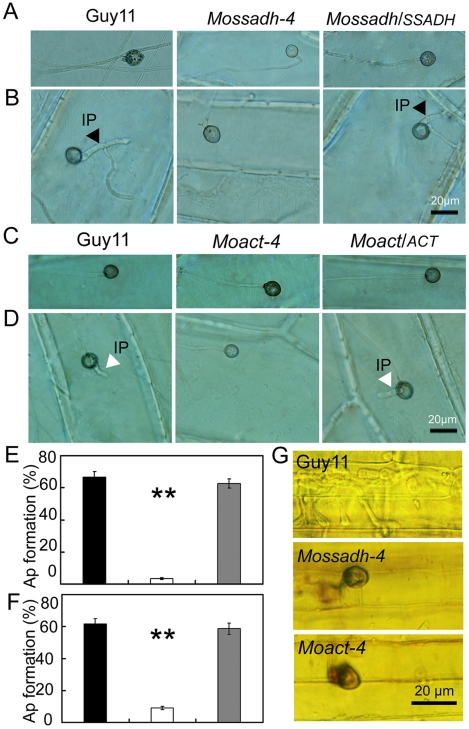
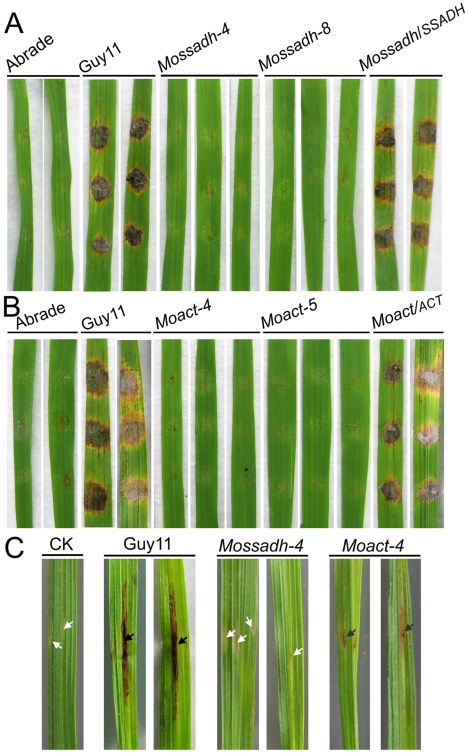
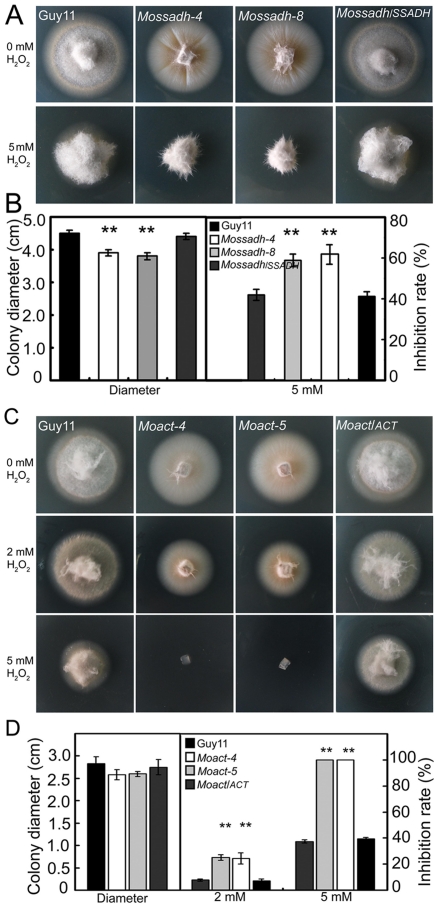
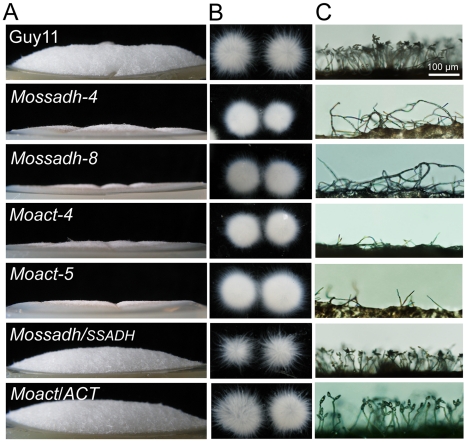
Saccharomyces cerevisiae Yap1 protein is an AP1-like transcription factor involved in the regulation of the oxidative stress response. An ortholog of Yap1, MoAP1, was recently identified from the rice blast fungus Magnaporthe oryzae genome. We found that MoAP1 is highly expressed in conidia and during invasive hyphal growth. The Moap1 mutant was sensitive to H₂O₂, similar to S. cerevisiae yap1 mutants, and MoAP1 complemented Yap1 function in resistance to H₂O₂, albeit partially. The Moap1 mutant also exhibited various defects in aerial hyphal growth, mycelial branching, conidia formation, the production of extracellular peroxidases and laccases, and melanin pigmentation. Consequently, the Moap1 mutant was unable to infect the host plant. The MoAP1-eGFP fusion protein is localized inside the nucleus upon exposure to H₂O₂, suggesting that MoAP1 also functions as a redox sensor. Moreover, through RNA sequence analysis, many MoAP1-regulated genes were identified, including several novel ones that were also involved in pathogenicity. Disruption of respective MGG_01662 (MoAAT) and MGG_02531 (encoding hypothetical protein) genes did not result in any detectable changes in conidial germination and appressorium formation but reduced pathogenicity, whereas the mutant strains of MGG_01230 (MoSSADH) and MGG_15157 (MoACT) showed marketed reductions in aerial hyphal growth, mycelial branching, and loss of conidiation as well as pathogenicity, similar to the Moap1 mutant. Taken together, our studies identify MoAP1 as a positive transcription factor that regulates transcriptions of MGG_01662, MGG_02531, MGG_01230, and MGG_15157 that are important in the growth, development, and pathogenicity of M. oryzae.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Doke N, Miura Y, Sanchez LM, Park HJ, Noritake T, et al. The oxidative burst protects plants against pathogen attack: mechanism and role as an emergency signal for plant bio-defence-a review. Gene. 1996;179:45–51. - PubMed
-
- Chen SX, Schopfer P. Hydroxyl-radical production in physiological reactions. A novel function of peroxidase. Eur J Biochem. 1999;260:726–735. - PubMed
-
- Torres MA, Dangl JL. Functions of the respiratory burst oxidase in biotic interactions, abiotic stress and development. Curr Opin Plant Biol. 2005;8:397–403. - PubMed
-
- Nurnberger T, Brunner F, Kemmerling B, Piater L. Innate immunity in plants and animals: striking similarities and obvious differences. Immunol Rev. 2004;198:249–266. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
