

Model for the exceptional reactivity of peroxiredoxins 2 and 3 with hydrogen peroxide: a kinetic and computational study
- PMID: 21385867
- PMCID: PMC3093878
- DOI: 10.1074/jbc.M111.232355
Model for the exceptional reactivity of peroxiredoxins 2 and 3 with hydrogen peroxide: a kinetic and computational study
Abstract
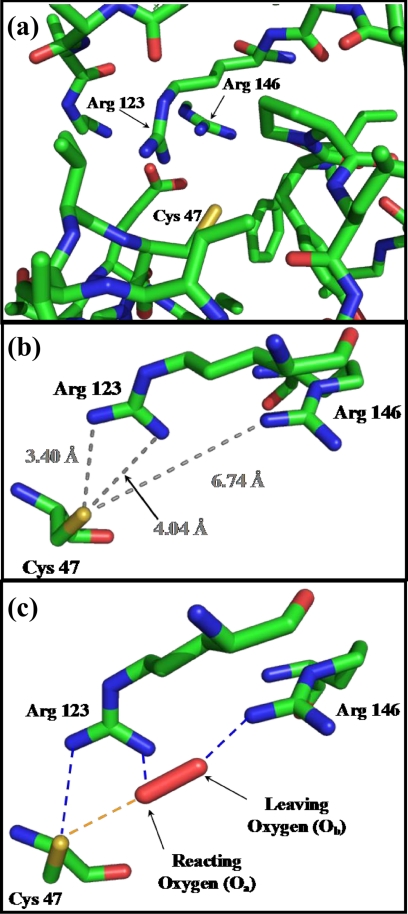
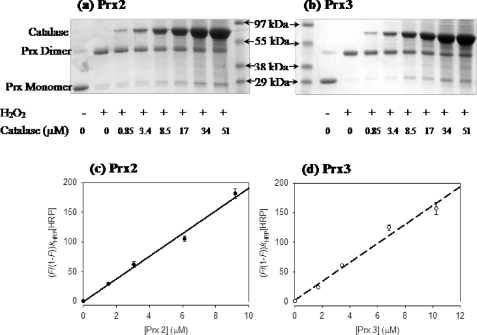
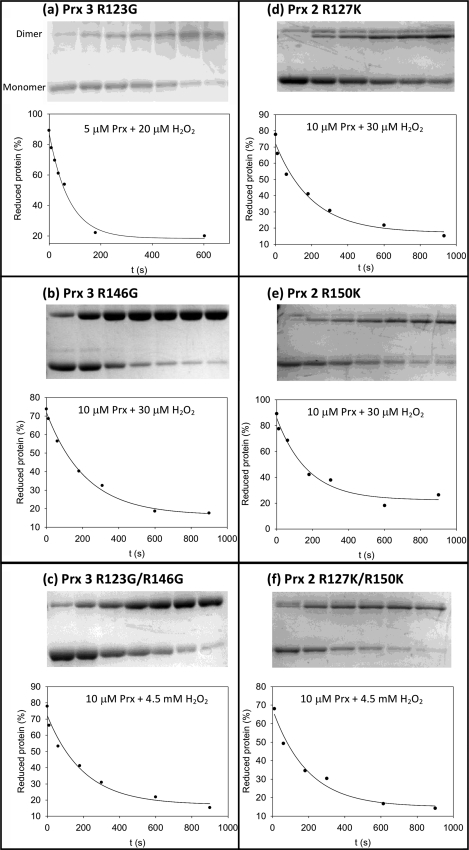
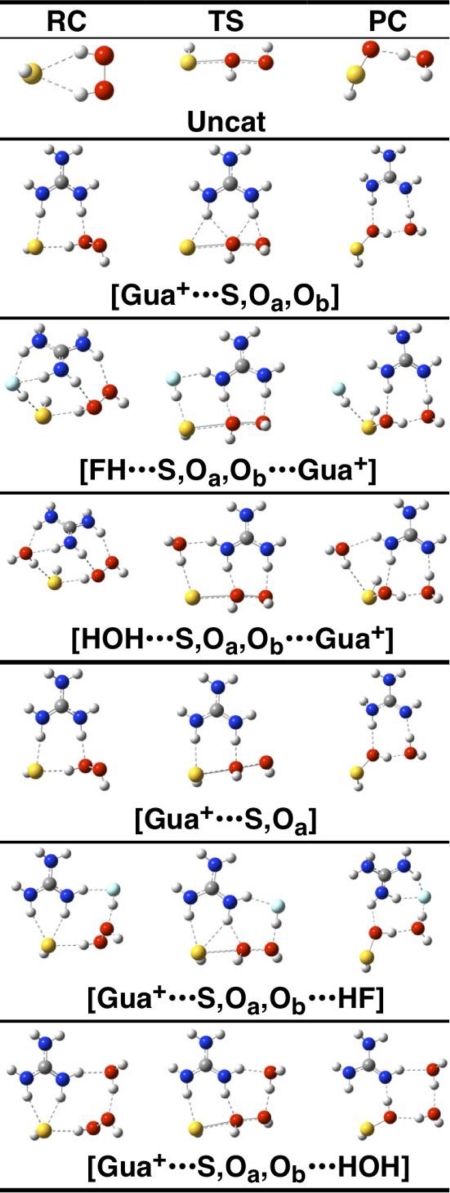
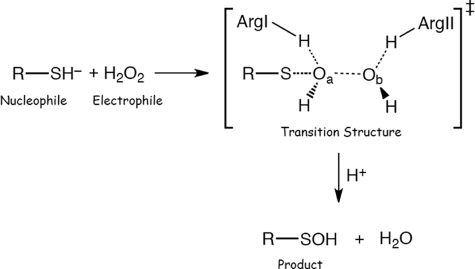
Peroxiredoxins (Prx) are thiol peroxidases that exhibit exceptionally high reactivity toward peroxides, but the chemical basis for this is not well understood. We present strong experimental evidence that two highly conserved arginine residues play a vital role in this activity of human Prx2 and Prx3. Point mutation of either ArgI or ArgII (in Prx3 Arg-123 and Arg-146, which are ∼3-4 Å or ∼6-7 Å away from the active site peroxidative cysteine (C(p)), respectively) in each case resulted in a 5 orders of magnitude loss in reactivity. A further 2 orders of magnitude decrease in the second-order rate constant was observed for the double arginine mutants of both isoforms, suggesting a cooperative function for these residues. Detailed ab initio theoretical calculations carried out with the high level G4 procedure suggest strong catalytic effects of H-bond-donating functional groups to the C(p) sulfur and the reactive and leaving oxygens of the peroxide in a cooperative manner. Using a guanidinium cation in the calculations to mimic the functional group of arginine, we were able to locate two transition structures that indicate rate enhancements consistent with our experimentally observed rate constants. Our results provide strong evidence for a vital role of ArgI in activating the peroxide that also involves H-bonding to ArgII. This mechanism could explain the exceptional reactivity of peroxiredoxins toward H(2)O(2) and may have wider implications for protein thiol reactivity toward peroxides.
© 2011 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Cox A. G., Winterbourn C. C., Hampton M. B. (2010) Biochem. J. 425, 313–325 - PubMed
-
- Fourquet S., Huang M. E., D'Autreaux B., Toledano M. B. (2008) Antioxid. Redox. Signal. 10, 1565–1576 - PubMed
-
- Cox A. G., Peskin A. V., Paton L. N., Winterbourn C. C., Hampton M. B. (2009) Biochemistry 48, 6495–6501 - PubMed
-
- Peskin A. V., Low F. M., Paton L. N., Maghzal G. J., Hampton M. B., Winterbourn C. C. (2007) J. Biol. Chem. 282, 11885–11892 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
