

Transcriptional and posttranslational up-regulation of HER3 (ErbB3) compensates for inhibition of the HER2 tyrosine kinase
- PMID: 21385943
- PMCID: PMC3064360
- DOI: 10.1073/pnas.1016140108
Transcriptional and posttranslational up-regulation of HER3 (ErbB3) compensates for inhibition of the HER2 tyrosine kinase
Abstract
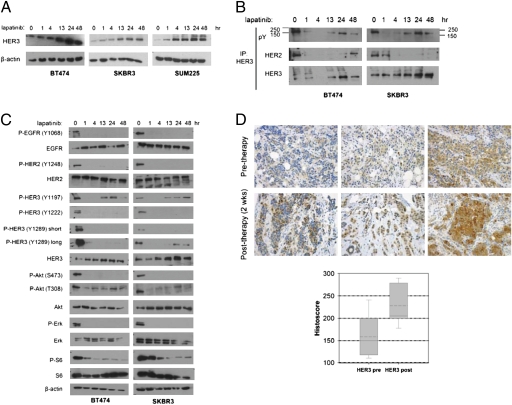
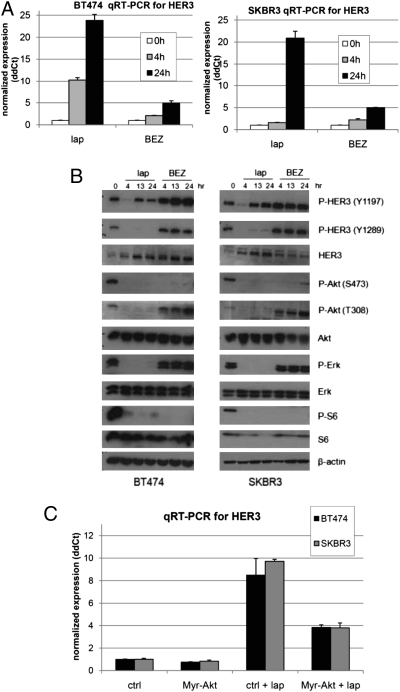
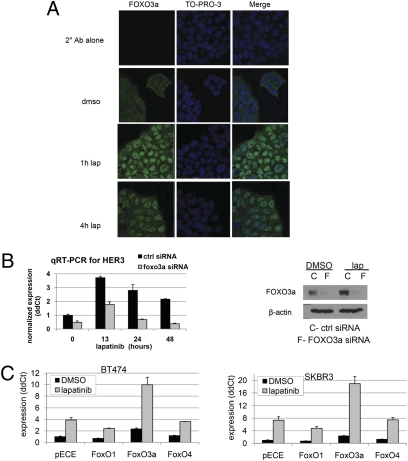
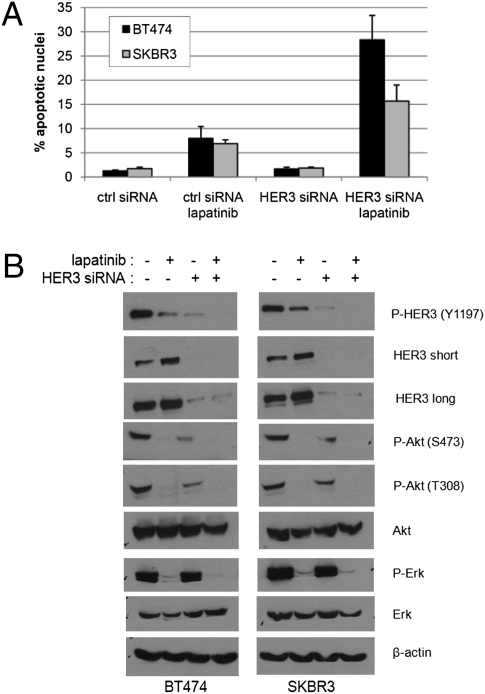
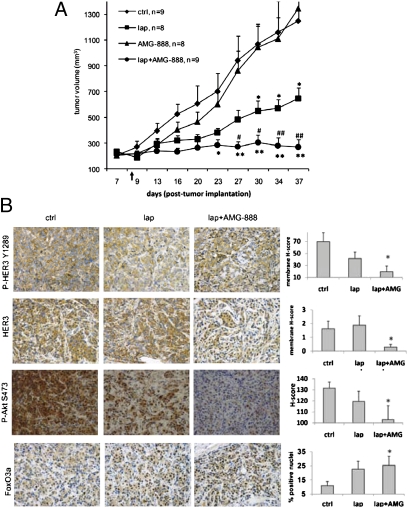
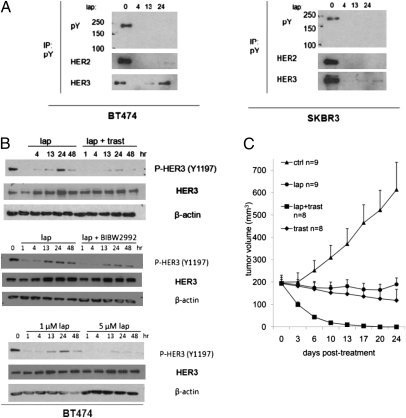
Sustained and complete inhibition of HER3 and its output to PI3K/Akt are required for the optimal antitumor effect of therapeutic inhibitors of the HER2 oncogene. Here, we show that, after inhibition of the HER2 tyrosine kinase with lapatinib, there is PI3K/Akt and FoxO3a-dependent up-regulation of HER3 mRNA and protein. Up-regulated HER3 was then phosphorylated by residual HER2 activity, thus partially maintaining P-Akt and limiting the antitumor action of lapatinib. Inhibition of HER3 with siRNA or a neutralizing HER3 antibody sensitized HER2+ breast cancer cells and xenografts to lapatinib both in vitro and in vivo. Combined blockade of HER2 and HER3 inhibited pharmacodynamic biomarkers of PI3K/Akt activity more effectively than each inhibitor alone. These results suggest that because of HER3-mediated compensation, current clinical inhibitors of HER2 and PI3K/Akt will not block the PI3K pathway completely. They also suggest that therapeutic inhibitors of HER3 should be used in combination with HER2 inhibitors and PI3K pathway inhibitors in patients with HER2- and PI3K-dependent cancers.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ross JS, Fletcher JA. The HER-2/neu oncogene in breast cancer: Prognostic factor, predictive factor, and target for therapy. Stem Cells. 1998;16:413–428. - PubMed
-
- Slamon DJ, et al. Human breast cancer: Correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science. 1987;235:177–182. - PubMed
-
- Alimandi M, et al. Cooperative signaling of ErbB3 and ErbB2 in neoplastic transformation and human mammary carcinomas. Oncogene. 1995;10:1813–1821. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
