

Second messengers and membrane trafficking direct and organize growth cone steering
- PMID: 21386859
- PMCID: PMC3133775
- DOI: 10.1038/nrn2996
Second messengers and membrane trafficking direct and organize growth cone steering
Abstract
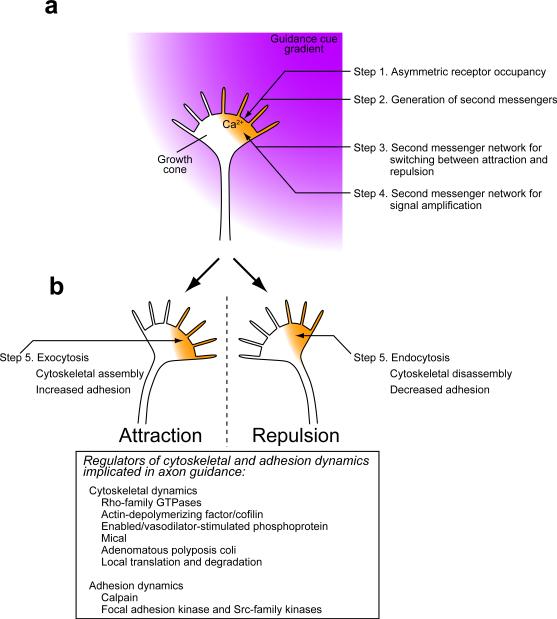
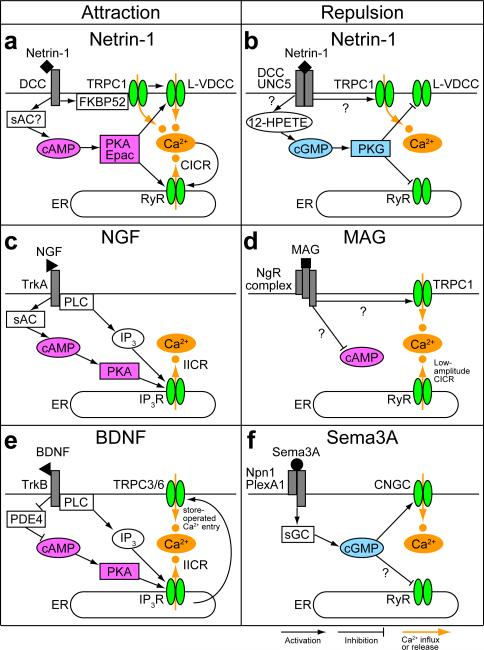
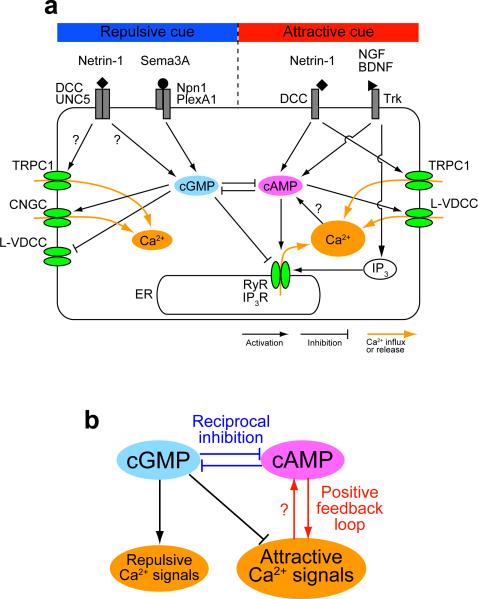
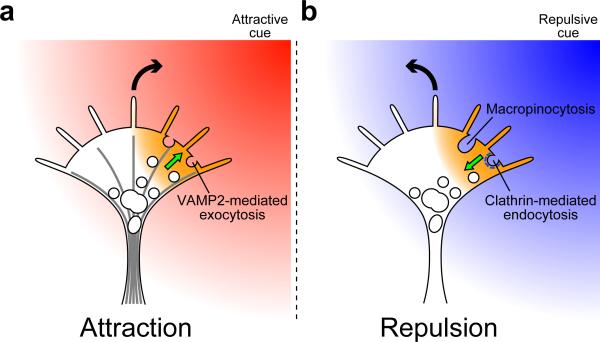
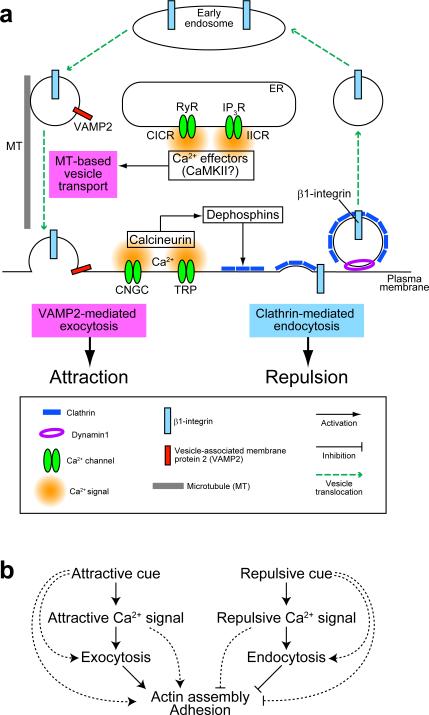
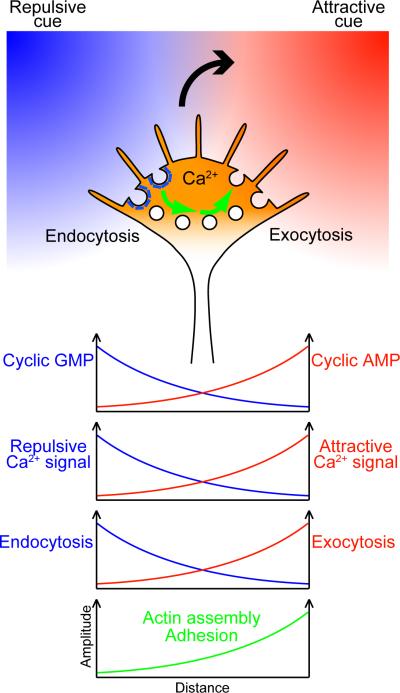
Graded distributions of extracellular cues guide developing axons toward their targets. A network of second messengers - Ca(2+) and cyclic nucleotides - shapes cue-derived information into either attractive or repulsive signals that steer growth cones bidirectionally. Emerging evidence suggests that such guidance signals create a localized imbalance between exocytosis and endocytosis, which in turn redirects membrane, adhesion and cytoskeletal components asymmetrically across the growth cone to bias the direction of axon extension. These recent advances allow us to propose a unifying model of how the growth cone translates shallow gradients of environmental information into polarized activity of the steering machinery for axon guidance.
Figures
References
-
- Sanes JR, Yamagata M. Many paths to synaptic specificity. Annu Rev Cell Dev Biol. 2009;25:161–195. - PubMed
-
- Sakano H. Neural map formation in the mouse olfactory system. Neuron. 2010;67:530–542. - PubMed
-
- Flanagan JG. Neural map specification by gradients. Curr Opin Neurobiol. 2006;16:59–66. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
