

The Populus Class III HD ZIP transcription factor POPCORONA affects cell differentiation during secondary growth of woody stems
- PMID: 21386988
- PMCID: PMC3046250
- DOI: 10.1371/journal.pone.0017458
The Populus Class III HD ZIP transcription factor POPCORONA affects cell differentiation during secondary growth of woody stems
Abstract
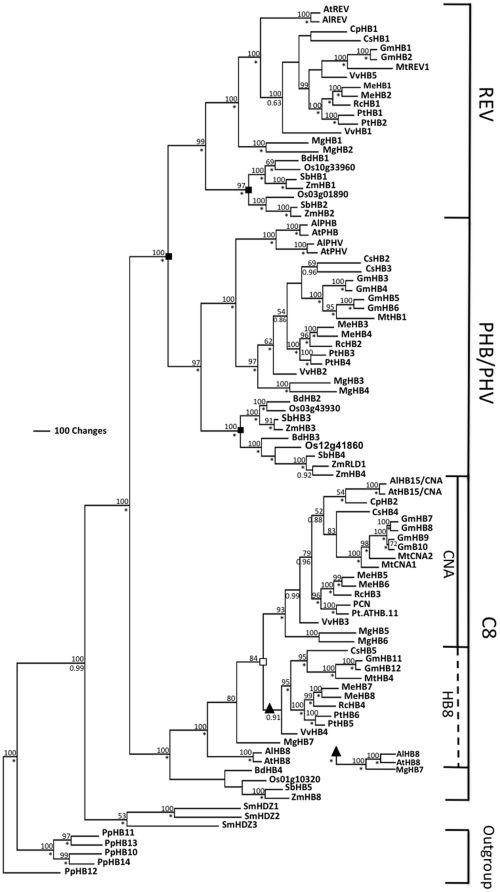
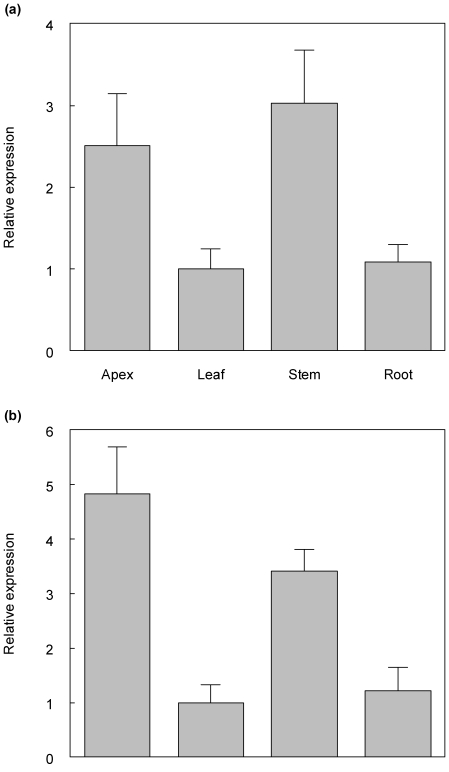
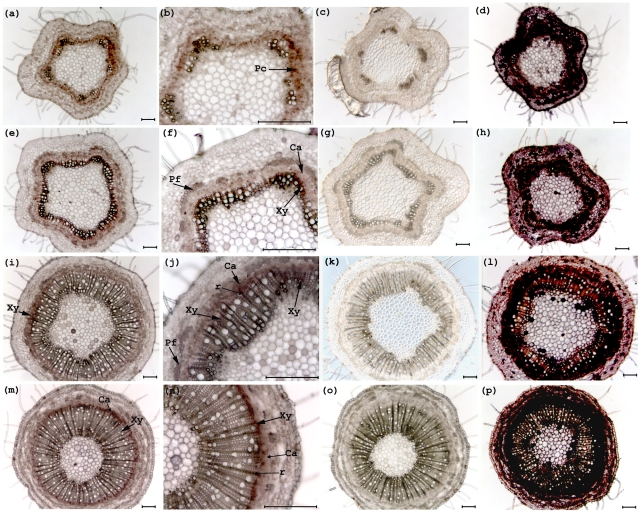
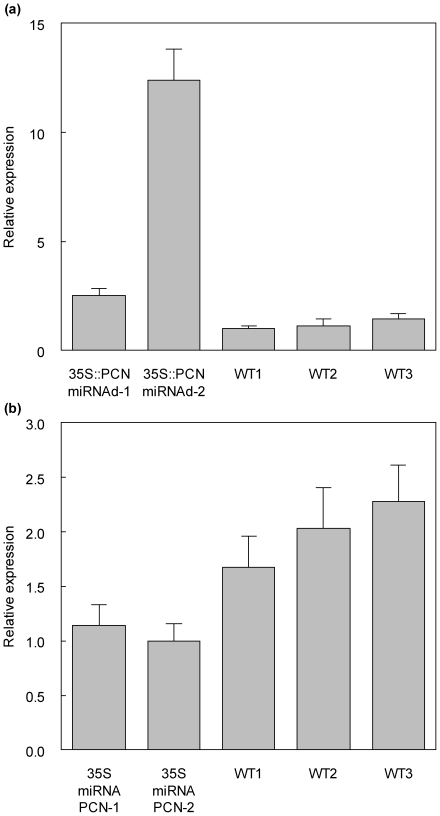
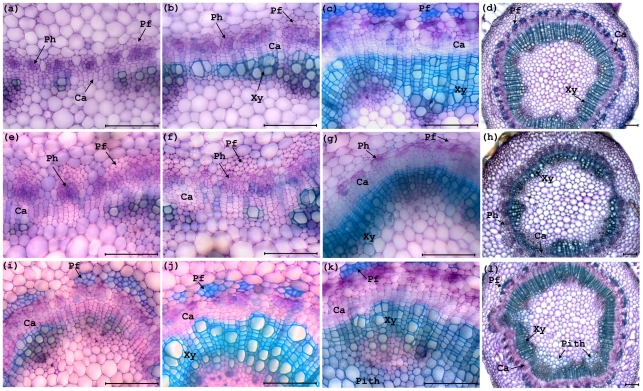
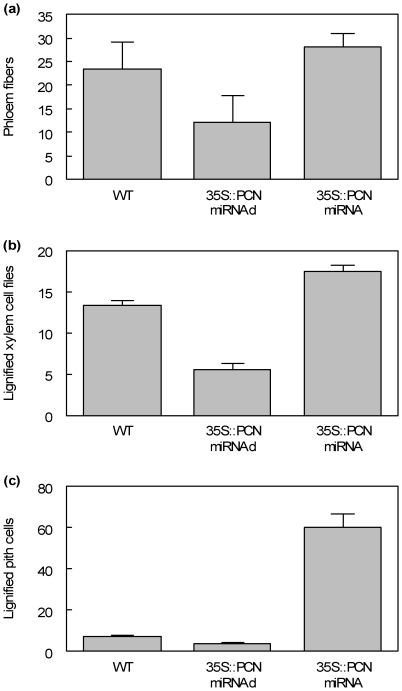
The developmental mechanisms regulating cell differentiation and patterning during the secondary growth of woody tissues are poorly understood. Class III HD ZIP transcription factors are evolutionarily ancient and play fundamental roles in various aspects of plant development. Here we investigate the role of a Class III HD ZIP transcription factor, POPCORONA, during secondary growth of woody stems. Transgenic Populus (poplar) trees expressing either a miRNA-resistant POPCORONA or a synthetic miRNA targeting POPCORONA were used to infer function of POPCORONA during secondary growth. Whole plant, histological, and gene expression changes were compared for transgenic and wild-type control plants. Synthetic miRNA knock down of POPCORONA results in abnormal lignification in cells of the pith, while overexpression of a miRNA-resistant POPCORONA results in delayed lignification of xylem and phloem fibers during secondary growth. POPCORONA misexpression also results in coordinated changes in expression of genes within a previously described transcriptional network regulating cell differentiation and cell wall biosynthesis, and hormone-related genes associated with fiber differentiation. POPCORONA illustrates another function of Class III HD ZIPs: regulating cell differentiation during secondary growth.
Conflict of interest statement
Figures

References
-
- Groover A, Nieminen K, Helariutta Y, Mansfield SD. Wood formation in Populus. 2010. in, Genetics and Genomics of Populus Jansson, Bhalerao, and Groover (eds) Springer.
-
- Larson PR. The Vascular Cambium. 1994. Springer-Verlag.
-
- Groover AT. What genes make a tree a tree? Trends in Plant Science. 2005;10:210–214. - PubMed
-
- Spicer R, Groover A. The evolution of development of the vascular cambium and secondary growth. New Phytol. 2010;186:577–592. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
