

Biogenic volatile organic compound and respiratory CO2 emissions after 13C-labeling: online tracing of C translocation dynamics in poplar plants
- PMID: 21387007
- PMCID: PMC3046154
- DOI: 10.1371/journal.pone.0017393
Biogenic volatile organic compound and respiratory CO2 emissions after 13C-labeling: online tracing of C translocation dynamics in poplar plants
Abstract
Background: Globally plants are the primary sink of atmospheric CO(2), but are also the major contributor of a large spectrum of atmospheric reactive hydrocarbons such as terpenes (e.g. isoprene) and other biogenic volatile organic compounds (BVOC). The prediction of plant carbon (C) uptake and atmospheric oxidation capacity are crucial to define the trajectory and consequences of global environmental changes. To achieve this, the biosynthesis of BVOC and the dynamics of C allocation and translocation in both plants and ecosystems are important.
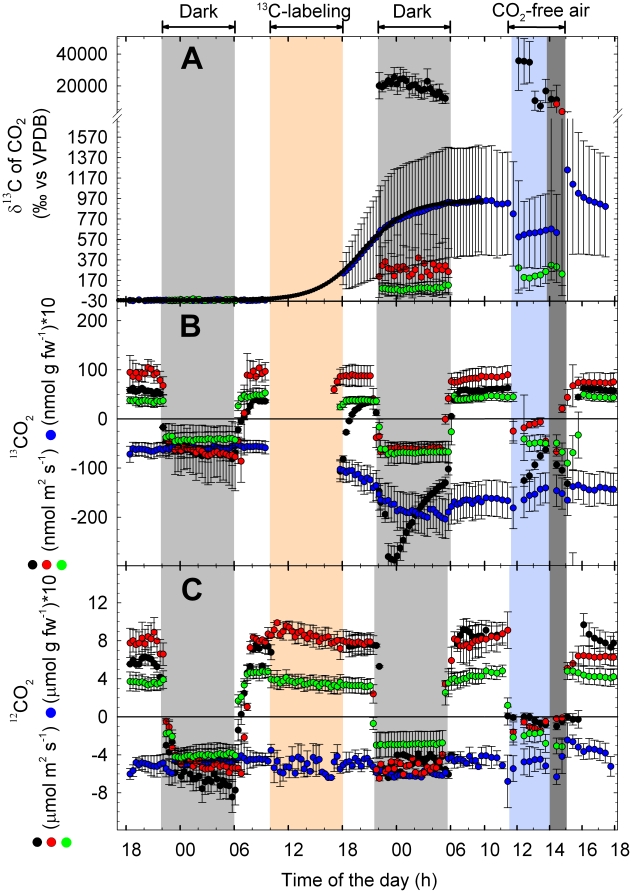
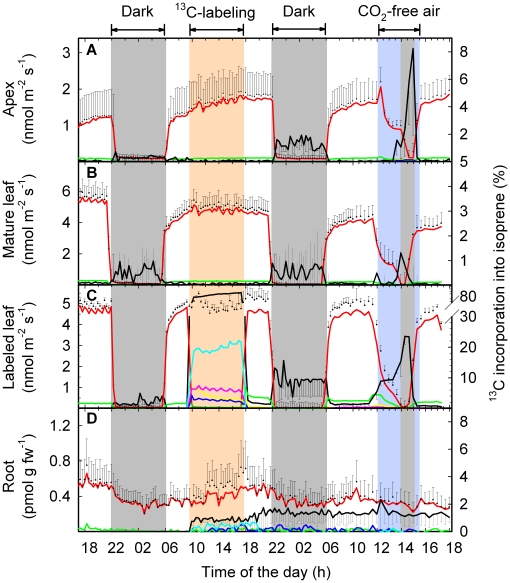
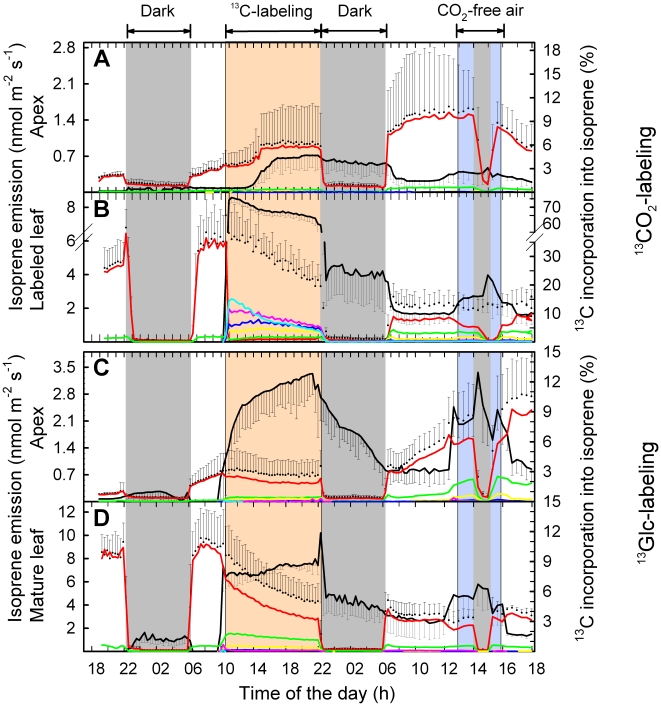
Methodology: We combined tunable diode laser absorption spectrometry (TDLAS) and proton transfer reaction mass spectrometry (PTR-MS) for studying isoprene biosynthesis and following C fluxes within grey poplar (Populus x canescens) saplings. This was achieved by feeding either (13)CO(2) to leaves or (13)C-glucose to shoots via xylem uptake. The translocation of (13)CO(2) from the source to other plant parts could be traced by (13)C-labeled isoprene and respiratory (13)CO(2) emission.
Principal finding: In intact plants, assimilated (13)CO(2) was rapidly translocated via the phloem to the roots within 1 hour, with an average phloem transport velocity of 20.3±2.5 cm h(-1). (13)C label was stored in the roots and partially reallocated to the plants' apical part one day after labeling, particularly in the absence of photosynthesis. The daily C loss as BVOC ranged between 1.6% in mature leaves and 7.0% in young leaves. Non-isoprene BVOC accounted under light conditions for half of the BVOC C loss in young leaves and one-third in mature leaves. The C loss as isoprene originated mainly (76-78%) from recently fixed CO(2), to a minor extent from xylem-transported sugars (7-11%) and from photosynthetic intermediates with slower turnover rates (8-11%).
Conclusion: We quantified the plants' C loss as respiratory CO(2) and BVOC emissions, allowing in tandem with metabolic analysis to deepen our understanding of ecosystem C flux.
Conflict of interest statement
Figures







References
-
- Prentice IC, Farquhar GD, Fasham MJR, Goulden ML, Heimann M, et al. The carbon cycle and atmospheric carbon dioxide. In: Houghton JT, Ding Y, Griggs DJ, Noguer M, van der Linden PJ, et al., editors. Climate Change 2001: The Physical Science Basis. Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge and New York, NY, UKUSA: Cambridge University Press; 2001. pp. 183–238.
-
- Forster P, Ramaswamy V, Artaxo P, Berntsen T, Betts R, et al. Changes in atmospheric constituents and in radiative forcing. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, et al., editors. Climate Change. Vol. 2007. CambridgeNew York, NY, United KingdomUSA: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press; 2007. pp. 129–234.
-
- Trainer M, Hsie E, McKeen S, Tallamraju R, Parrish D, et al. Impact of natural hydrocarbons on hydroxyl and peroxy radicals at a remote site. J Geophys Res. 1987;92:11879–11894.
-
- Chameides WL, Lindsay RW, Richardson J, Kiang CS. The role of biogenic hydrocarbons in urban photochemical smog: Atlanta as a case study. Science. 1988;241:1473–1475. - PubMed
-
- Jacob DJ, Wofsy SC. Photochemistry of biogenic emissions over the Amazon Forest. J Geophys Res-Atmos. 1988;93:1477–1486.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
