

Bub1, Sgo1, and Mps1 mediate a distinct pathway for chromosome biorientation in budding yeast
- PMID: 21389114
- PMCID: PMC3084670
- DOI: 10.1091/mbc.E10-08-0673
Bub1, Sgo1, and Mps1 mediate a distinct pathway for chromosome biorientation in budding yeast
Abstract
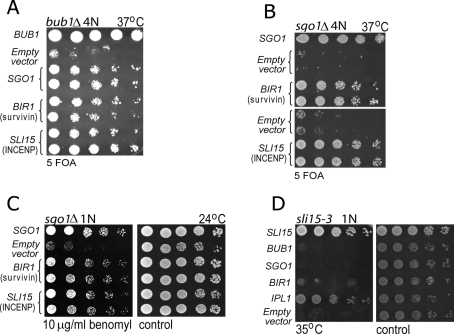
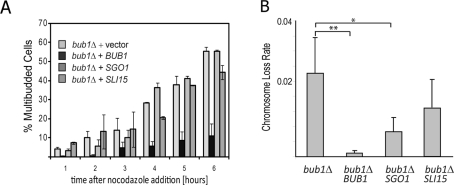
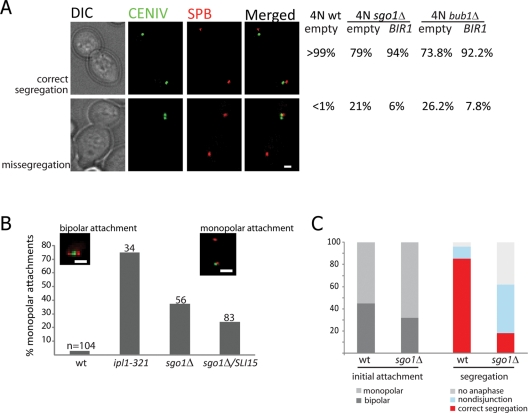
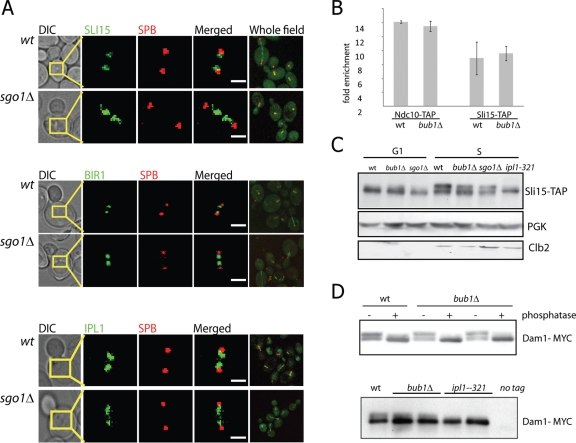
The conserved mitotic kinase Bub1 performs multiple functions that are only partially characterized. Besides its role in the spindle assembly checkpoint and chromosome alignment, Bub1 is crucial for the kinetochore recruitment of multiple proteins, among them Sgo1. Both Bub1 and Sgo1 are dispensable for growth of haploid and diploid budding yeast, but they become essential in cells with higher ploidy. We find that overexpression of SGO1 partially corrects the chromosome segregation defect of bub1Δ haploid cells and restores viability to bub1Δ tetraploid cells. Using an unbiased high-copy suppressor screen, we identified two members of the chromosomal passenger complex (CPC), BIR1 (survivin) and SLI15 (INCENP, inner centromere protein), as suppressors of the growth defect of both bub1Δ and sgo1Δ tetraploids, suggesting that these mutants die due to defects in chromosome biorientation. Overexpression of BIR1 or SLI15 also complements the benomyl sensitivity of haploid bub1Δ and sgo1Δ cells. Mutants lacking SGO1 fail to biorient sister chromatids attached to the same spindle pole (syntelic attachment) after nocodazole treatment. Moreover, the sgo1Δ cells accumulate syntelic attachments in unperturbed mitoses, a defect that is partially corrected by BIR1 or SLI15 overexpression. We show that in budding yeast neither Bub1 nor Sgo1 is required for CPC localization or affects Aurora B activity. Instead we identify Sgo1 as a possible partner of Mps1, a mitotic kinase suggested to have an Aurora B-independent function in establishment of biorientation. We found that Sgo1 overexpression rescues defects caused by metaphase inactivation of Mps1 and that Mps1 is required for Sgo1 localization to the kinetochore. We propose that Bub1, Sgo1, and Mps1 facilitate chromosome biorientation independently of the Aurora B-mediated pathway at the budding yeast kinetochore and that both pathways are required for the efficient turnover of syntelic attachments.
Figures
Similar articles
-
Tension sensing by Aurora B kinase is independent of survivin-based centromere localization.Nature. 2013 May 2;497(7447):118-21. doi: 10.1038/nature12057. Epub 2013 Apr 21. Nature. 2013. PMID: 23604256 Free PMC article.
-
Bub1 kinase targets Sgo1 to ensure efficient chromosome biorientation in budding yeast mitosis.PLoS Genet. 2007 Nov;3(11):e213. doi: 10.1371/journal.pgen.0030213. Epub 2007 Oct 15. PLoS Genet. 2007. PMID: 18081426 Free PMC article.
-
Killing two birds with one stone: how budding yeast Mps1 controls chromosome segregation and spindle assembly checkpoint through phosphorylation of a single kinetochore protein.Curr Genet. 2020 Dec;66(6):1037-1044. doi: 10.1007/s00294-020-01091-x. Epub 2020 Jul 6. Curr Genet. 2020. PMID: 32632756 Review.
-
Efficient chromosome biorientation and the tension checkpoint in Saccharomyces cerevisiae both require Bir1.Mol Cell Biol. 2009 Aug;29(16):4552-62. doi: 10.1128/MCB.01911-08. Epub 2009 Jun 15. Mol Cell Biol. 2009. PMID: 19528231 Free PMC article.
-
The composition, functions, and regulation of the budding yeast kinetochore.Genetics. 2013 Aug;194(4):817-46. doi: 10.1534/genetics.112.145276. Genetics. 2013. PMID: 23908374 Free PMC article. Review.
Cited by
-
Genetic interactions between specific chromosome copy number alterations dictate complex aneuploidy patterns.Genes Dev. 2018 Dec 1;32(23-24):1485-1498. doi: 10.1101/gad.319400.118. Epub 2018 Nov 21. Genes Dev. 2018. PMID: 30463904 Free PMC article.
-
Genetic instability in budding and fission yeast-sources and mechanisms.FEMS Microbiol Rev. 2015 Nov;39(6):917-67. doi: 10.1093/femsre/fuv028. Epub 2015 Jun 24. FEMS Microbiol Rev. 2015. PMID: 26109598 Free PMC article. Review.
-
An engineered minimal chromosomal passenger complex reveals a role for INCENP/Sli15 spindle association in chromosome biorientation.J Cell Biol. 2017 Apr 3;216(4):911-923. doi: 10.1083/jcb.201609123. Epub 2017 Mar 17. J Cell Biol. 2017. PMID: 28314741 Free PMC article.
-
Tip60 acetylation of histone H3K4 temporally controls chromosome passenger complex localization.Mol Biol Cell. 2022 Aug 1;33(9):br15. doi: 10.1091/mbc.E21-06-0283. Epub 2022 Jun 2. Mol Biol Cell. 2022. PMID: 35653296 Free PMC article.
-
Mitotic kinases as drivers of the epithelial-to-mesenchymal transition and as therapeutic targets against breast cancers.Exp Biol Med (Maywood). 2021 May;246(9):1036-1044. doi: 10.1177/1535370221991094. Epub 2021 Feb 18. Exp Biol Med (Maywood). 2021. PMID: 33601912 Free PMC article. Review.
References
-
- Cheeseman IM, Anderson S, Jwa M, Green EM, Kang J, Yates JR III, Chan CS, Drubin DG, Barnes G. Phospho-regulation of kinetochore-microtubule attachments by the Aurora kinase Ipl1p. Cell. 2002;111:163–172. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
