

Spatiotemporal properties of neuron response suppression in owl monkey primary somatosensory cortex when stimuli are presented to both hands
- PMID: 21389215
- PMCID: PMC3063385
- DOI: 10.1523/JNEUROSCI.4310-10.2011
Spatiotemporal properties of neuron response suppression in owl monkey primary somatosensory cortex when stimuli are presented to both hands
Abstract
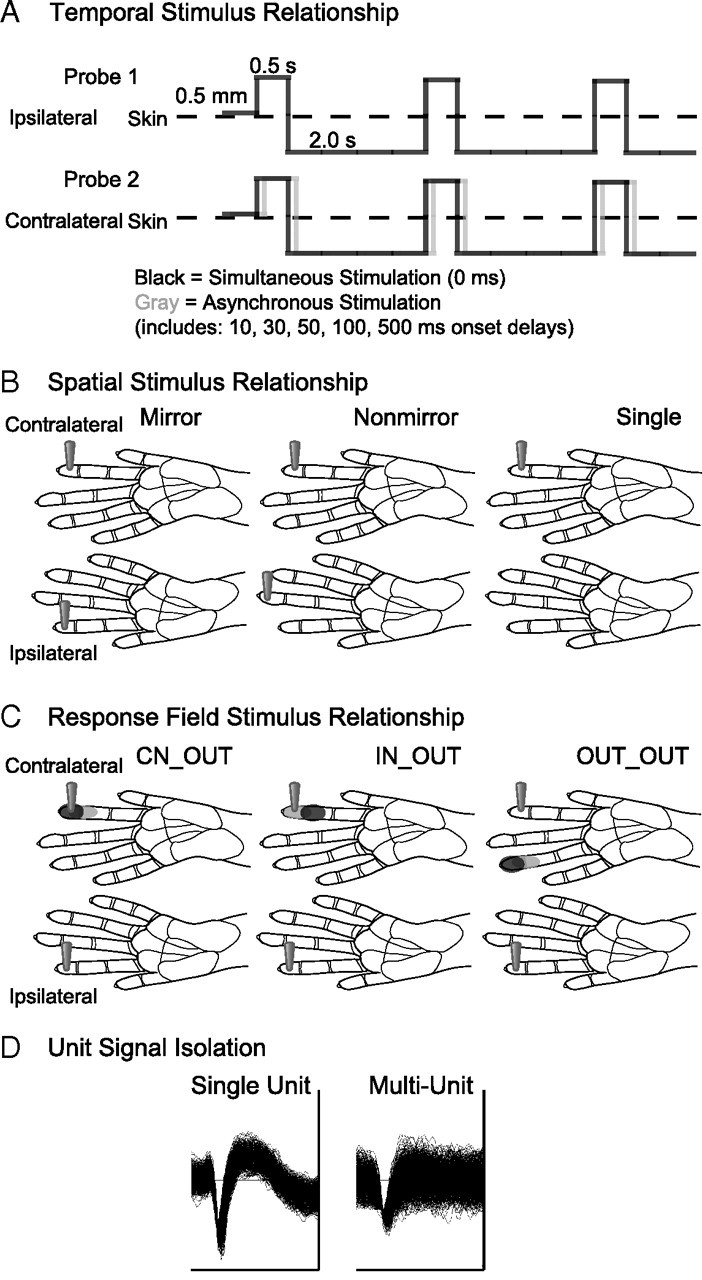
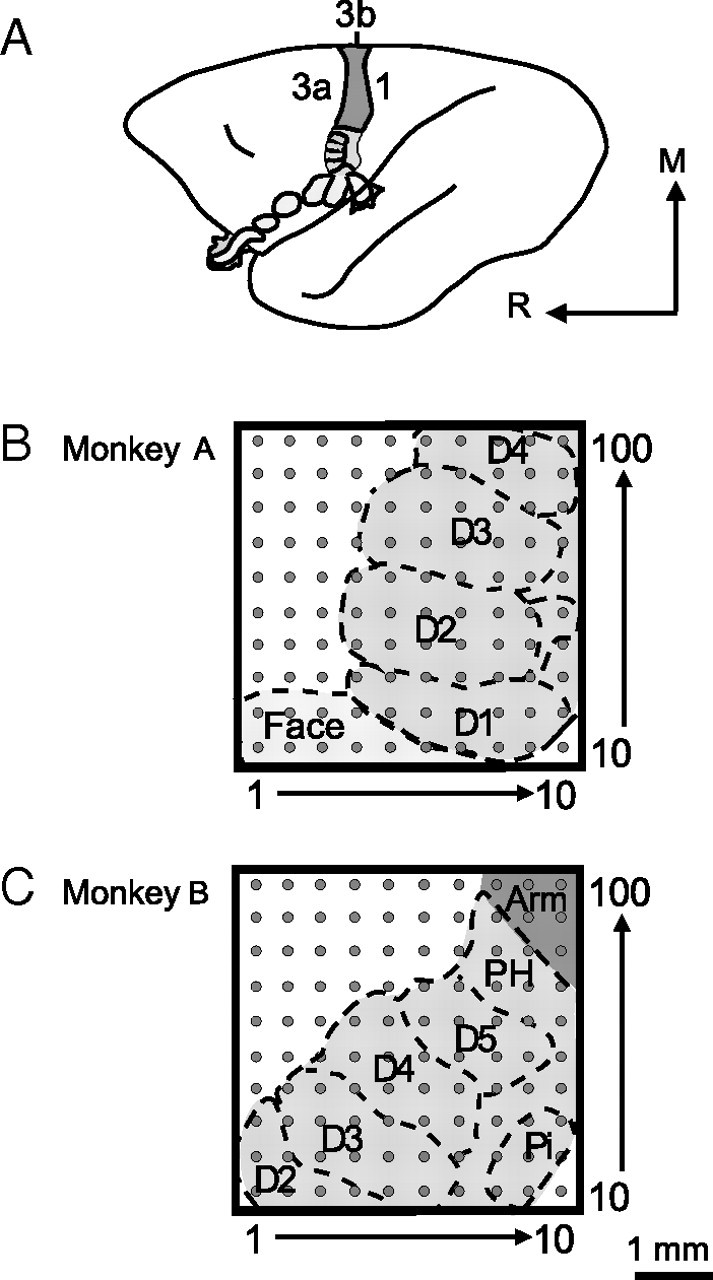
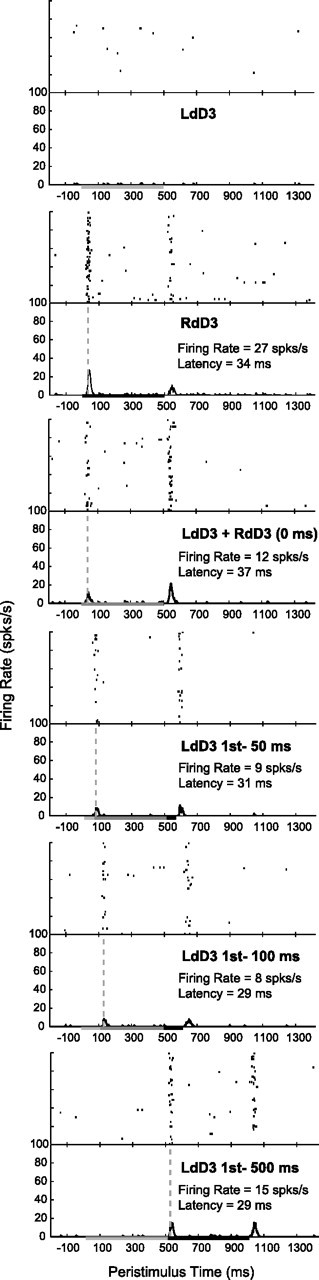
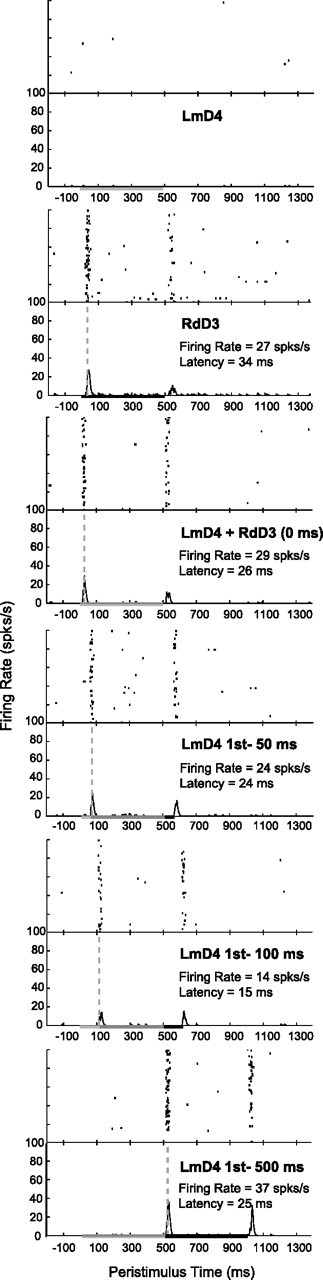
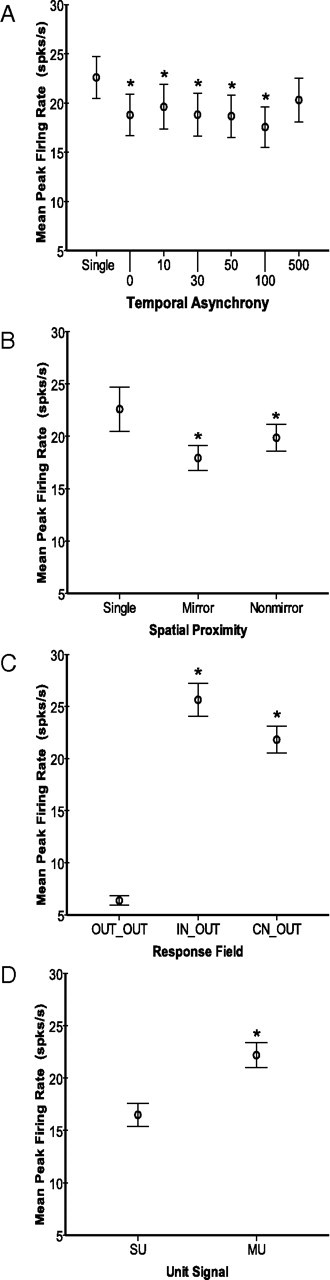
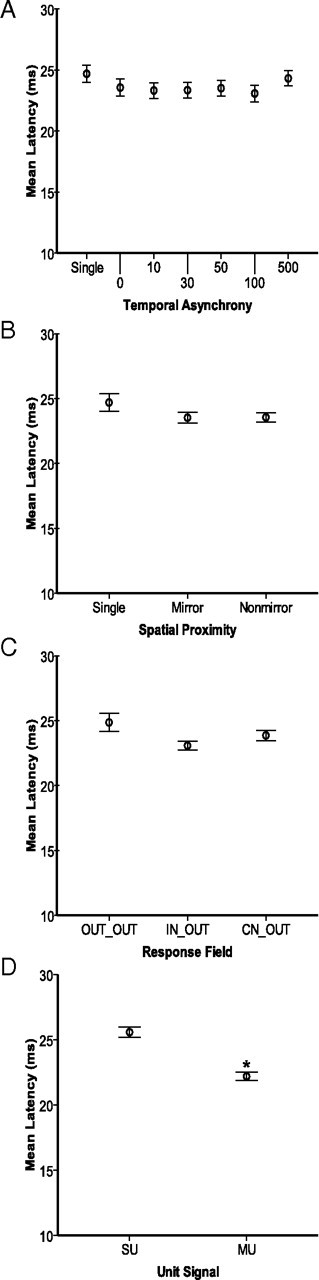
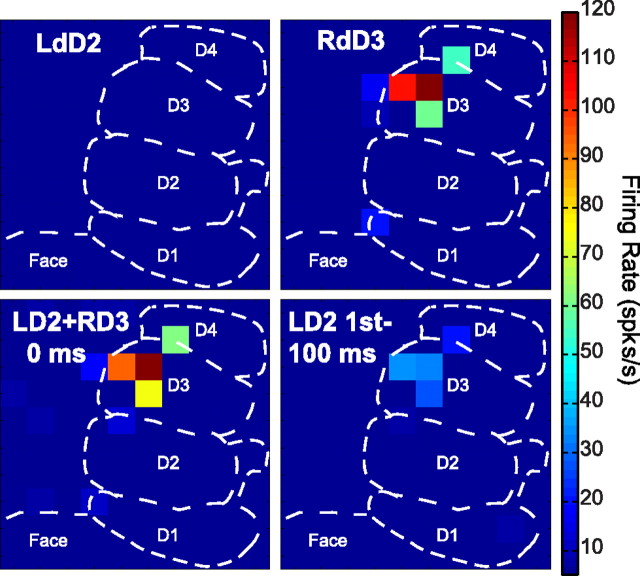
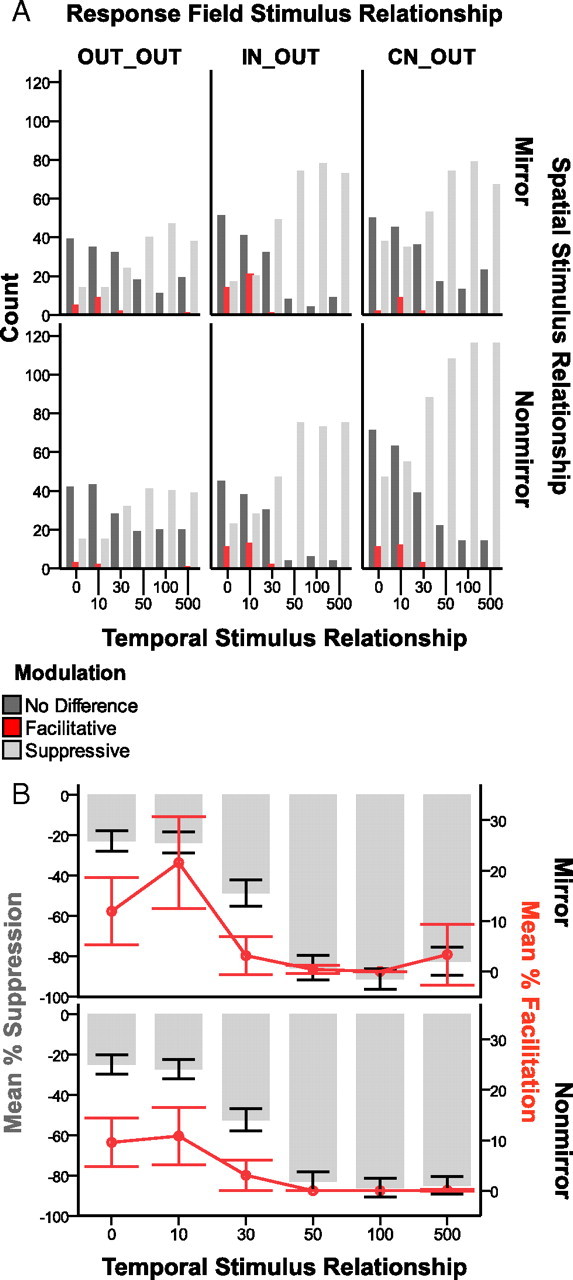
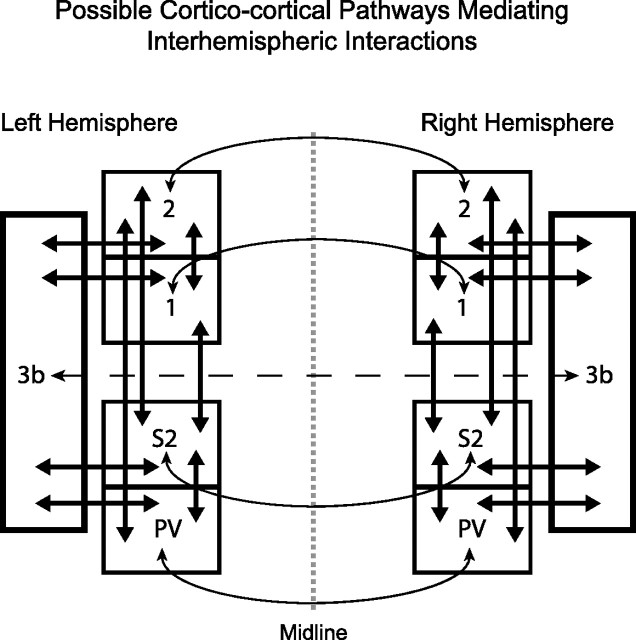
Despite the lack of ipsilateral receptive fields (RFs) for neurons in the hand representation of area 3b of primary somatosensory cortex, interhemispheric interactions have been reported to varying degrees. We investigated spatiotemporal properties of these interactions to determine the following: response types, timing between stimuli to evoke the strongest bimanual interactions, topographical distribution of effects, and their dependence on similarity of stimulus locations on the two hands. We analyzed response magnitudes and latencies of single neurons and multineuron clusters recorded from 100-electrode arrays implanted in one hemisphere of each of two anesthetized owl monkeys. Skin indentations were delivered to the two hands simultaneously and asynchronously at mirror locations (matched sites on each hand) and nonmirror locations. Since multiple neurons were recorded simultaneously, stimuli on the contralateral hand could be within or outside of the classical RFs of any given neuron. For most neurons, stimulation on the ipsilateral hand suppressed responses to stimuli on the contralateral hand. Maximum suppression occurred when the ipsilateral stimulus was presented 100 ms before the contralateral stimulus onset (p < 0.0005). The longest stimulus onset delay tested (500 ms) allowed contralateral responses to recover to control levels (p = 0.428). Stimulation on mirror digits did not differ from stimulation on nonmirror locations (p = 1.000). These results indicate that interhemispheric interactions are common in area 3b, somewhat topographically diffuse, and maximal when the suppressing ipsilateral stimulus precedes the contralateral stimulus. Our findings point to a neurophysiological basis for "interference" effects found in human psychophysical studies of bimanual stimulation.
Figures
Similar articles
-
Effects of spatiotemporal stimulus properties on spike timing correlations in owl monkey primary somatosensory cortex.J Neurophysiol. 2012 Dec;108(12):3353-69. doi: 10.1152/jn.00414.2011. Epub 2012 Sep 26. J Neurophysiol. 2012. PMID: 23019003 Free PMC article.
-
Response properties of neurons in primary somatosensory cortex of owl monkeys reflect widespread spatiotemporal integration.J Neurophysiol. 2010 Apr;103(4):2139-57. doi: 10.1152/jn.00709.2009. Epub 2010 Feb 17. J Neurophysiol. 2010. PMID: 20164400 Free PMC article.
-
Changes in the distributed temporal response properties of SI cortical neurons reflect improvements in performance on a temporally based tactile discrimination task.J Neurophysiol. 1992 May;67(5):1071-91. doi: 10.1152/jn.1992.67.5.1071. J Neurophysiol. 1992. PMID: 1597698
-
Topographic reorganization of the hand representation in cortical area 3b owl monkeys trained in a frequency-discrimination task.J Neurophysiol. 1992 May;67(5):1031-56. doi: 10.1152/jn.1992.67.5.1031. J Neurophysiol. 1992. PMID: 1597696
-
Multielectrode Recordings in the Somatosensory System.In: Nicolelis MAL, editor. Methods for Neural Ensemble Recordings. 2nd edition. Boca Raton (FL): CRC Press/Taylor & Francis; 2008. Chapter 6. In: Nicolelis MAL, editor. Methods for Neural Ensemble Recordings. 2nd edition. Boca Raton (FL): CRC Press/Taylor & Francis; 2008. Chapter 6. PMID: 21204443 Free Books & Documents. Review.
Cited by
-
A Conceptual Model of Tactile Processing across Body Features of Size, Shape, Side, and Spatial Location.Front Psychol. 2019 Feb 26;10:291. doi: 10.3389/fpsyg.2019.00291. eCollection 2019. Front Psychol. 2019. PMID: 30863333 Free PMC article. Review.
-
Cortical connections of the functional domain for climbing or running in posterior parietal cortex of galagos.J Comp Neurol. 2021 Jul 1;529(10):2789-2812. doi: 10.1002/cne.25123. Epub 2021 Mar 4. J Comp Neurol. 2021. PMID: 33550608 Free PMC article.
-
Effects of muscimol inactivations of functional domains in motor, premotor, and posterior parietal cortex on complex movements evoked by electrical stimulation.J Neurophysiol. 2014 Mar;111(5):1100-19. doi: 10.1152/jn.00491.2013. Epub 2013 Dec 18. J Neurophysiol. 2014. PMID: 24353298 Free PMC article.
-
Effects of spatiotemporal stimulus properties on spike timing correlations in owl monkey primary somatosensory cortex.J Neurophysiol. 2012 Dec;108(12):3353-69. doi: 10.1152/jn.00414.2011. Epub 2012 Sep 26. J Neurophysiol. 2012. PMID: 23019003 Free PMC article.
-
Distinct fine-scale fMRI activation patterns of contra- and ipsilateral somatosensory areas 3b and 1 in humans.Hum Brain Mapp. 2014 Sep;35(9):4841-57. doi: 10.1002/hbm.22517. Epub 2014 Apr 1. Hum Brain Mapp. 2014. PMID: 24692215 Free PMC article.
References
-
- Alenda A, Nuñez A. Sensory-interference in rat primary somatosensory cortical neurons. Eur J Neurosci. 2004;19:766–770. - PubMed
-
- Alvarado JC, Vaughan JW, Stanford TR, Stein BE. Multisensory versus unisensory integration: contrasting modes in the superior colliculus. J Neurophysiol. 2007;97:3193–3205. - PubMed
-
- Angelucci A, Levitt JB, Lund JS. Anatomical origins of the classical receptive field and modulatory surround field of single neurons in macaque visual cortical area V1. In: Azmita EC, DeFelipe J, Jones EG, Rakic P, Ribak CE, editors. Progress in brain research, Chap 29. Vol. 136. Elsevier Science B.V; 2002. pp. 373–388. - PubMed
-
- Braun C, Hess H, Burkhardt M, Wühle A, Preissl H. The right hand knows what the left hand is feeling. Exp Brain Res. 2005;162:366–373. - PubMed
-
- Burton H, Fabri M. Ipsilateral intracortical connections of physiologically defined cutaneous representations in area 3b and 1 of macaque monkeys: projections in the vicinity of the central sulcus. J Comp Neurol. 1995;355:508–538. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources