

Amygdala circuitry mediating reversible and bidirectional control of anxiety
- PMID: 21389985
- PMCID: PMC3154022
- DOI: 10.1038/nature09820
Amygdala circuitry mediating reversible and bidirectional control of anxiety
Abstract
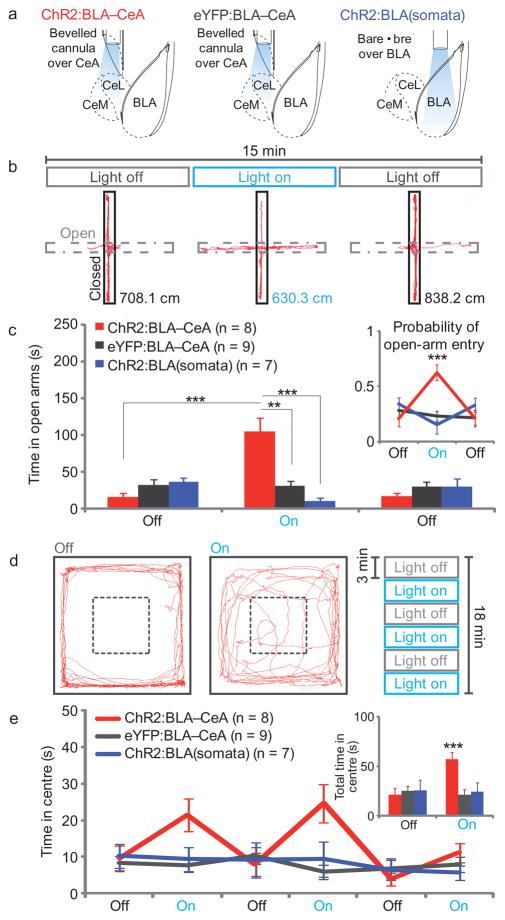
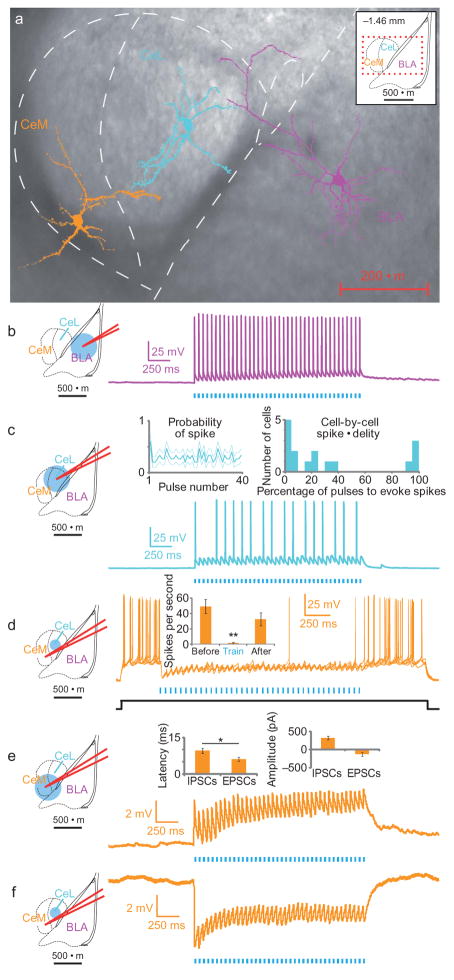
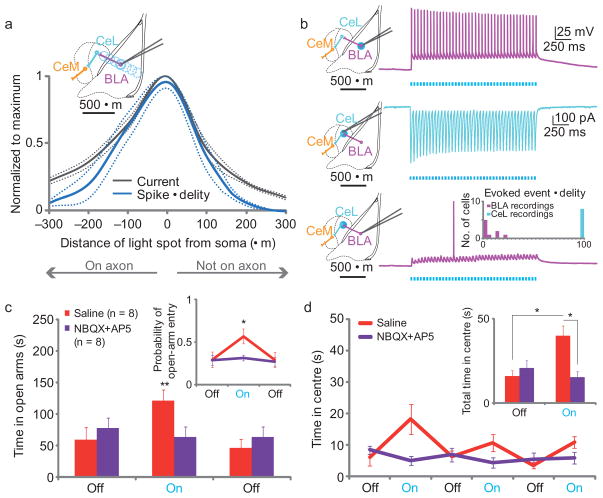
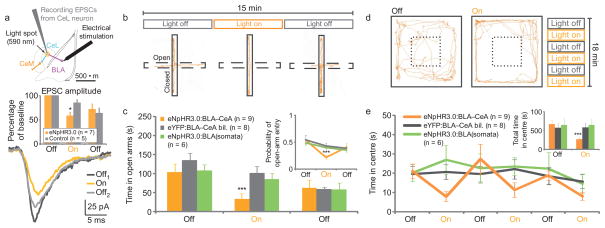
Anxiety--a sustained state of heightened apprehension in the absence of immediate threat--becomes severely debilitating in disease states. Anxiety disorders represent the most common of psychiatric diseases (28% lifetime prevalence) and contribute to the aetiology of major depression and substance abuse. Although it has been proposed that the amygdala, a brain region important for emotional processing, has a role in anxiety, the neural mechanisms that control anxiety remain unclear. Here we explore the neural circuits underlying anxiety-related behaviours by using optogenetics with two-photon microscopy, anxiety assays in freely moving mice, and electrophysiology. With the capability of optogenetics to control not only cell types but also specific connections between cells, we observed that temporally precise optogenetic stimulation of basolateral amygdala (BLA) terminals in the central nucleus of the amygdala (CeA)--achieved by viral transduction of the BLA with a codon-optimized channelrhodopsin followed by restricted illumination in the downstream CeA--exerted an acute, reversible anxiolytic effect. Conversely, selective optogenetic inhibition of the same projection with a third-generation halorhodopsin (eNpHR3.0) increased anxiety-related behaviours. Importantly, these effects were not observed with direct optogenetic control of BLA somata, possibly owing to recruitment of antagonistic downstream structures. Together, these results implicate specific BLA-CeA projections as critical circuit elements for acute anxiety control in the mammalian brain, and demonstrate the importance of optogenetically targeting defined projections, beyond simply targeting cell types, in the study of circuit function relevant to neuropsychiatric disease.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
