

DNA ligase III is critical for mtDNA integrity but not Xrcc1-mediated nuclear DNA repair
- PMID: 21390131
- PMCID: PMC3079429
- DOI: 10.1038/nature09773
DNA ligase III is critical for mtDNA integrity but not Xrcc1-mediated nuclear DNA repair
Abstract
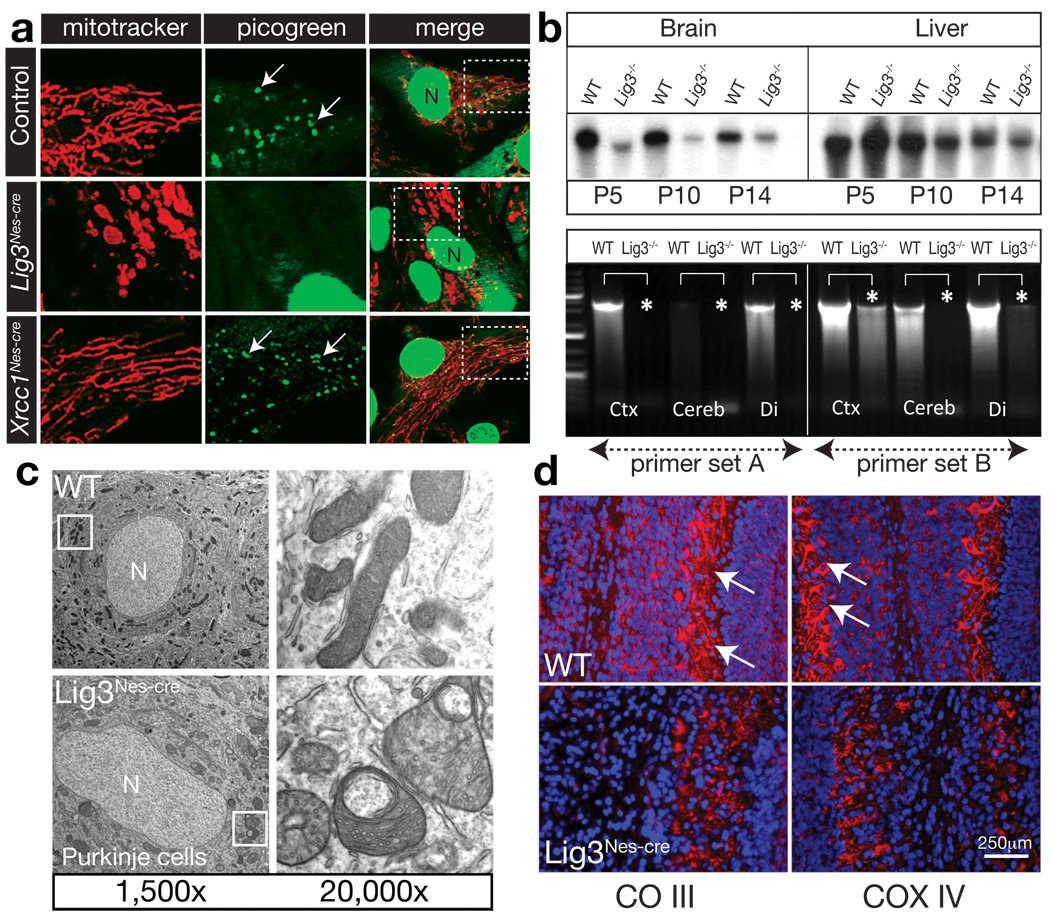
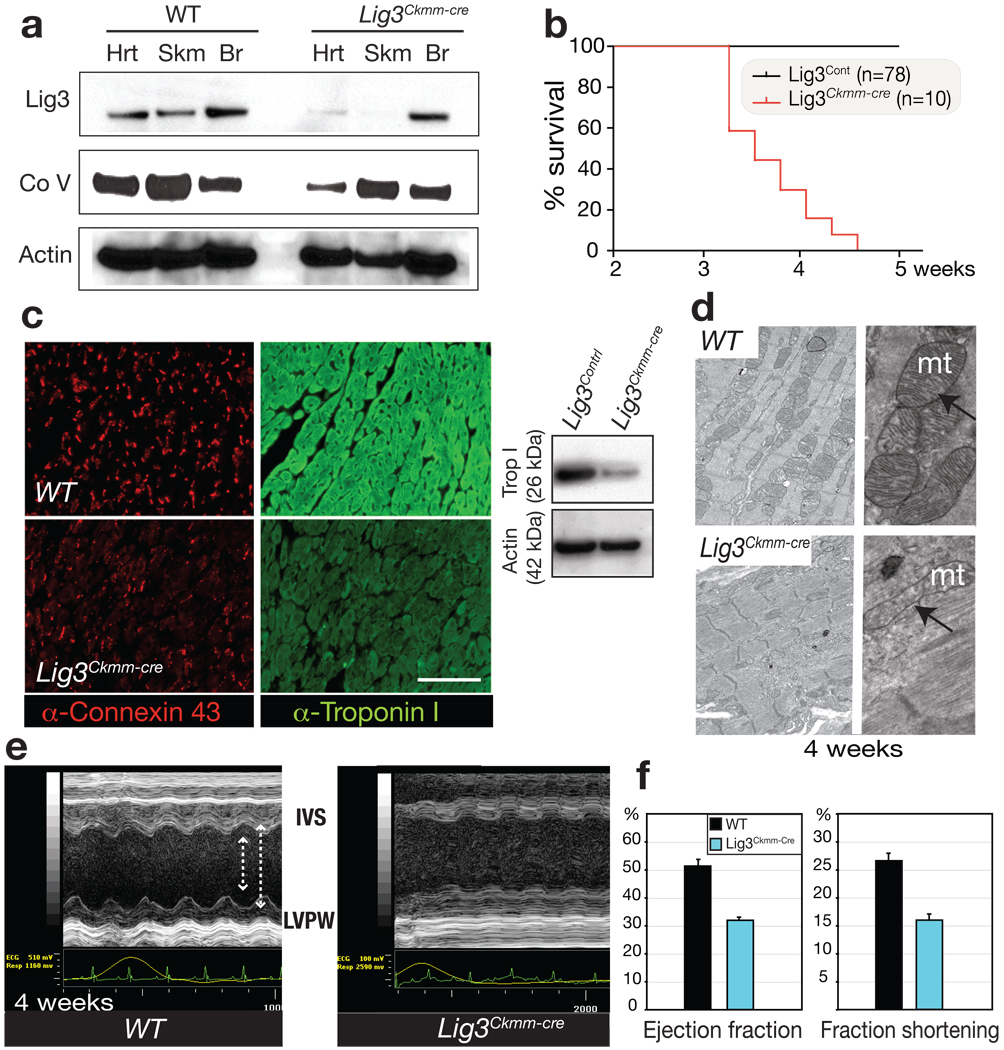
DNA replication and repair in mammalian cells involves three distinct DNA ligases: ligase I (Lig1), ligase III (Lig3) and ligase IV (Lig4). Lig3 is considered a key ligase during base excision repair because its stability depends upon its nuclear binding partner Xrcc1, a critical factor for this DNA repair pathway. Lig3 is also present in the mitochondria, where its role in mitochondrial DNA (mtDNA) maintenance is independent of Xrcc1 (ref. 4). However, the biological role of Lig3 is unclear as inactivation of murine Lig3 results in early embryonic lethality. Here we report that Lig3 is essential for mtDNA integrity but dispensable for nuclear DNA repair. Inactivation of Lig3 in the mouse nervous system resulted in mtDNA loss leading to profound mitochondrial dysfunction, disruption of cellular homeostasis and incapacitating ataxia. Similarly, inactivation of Lig3 in cardiac muscle resulted in mitochondrial dysfunction and defective heart-pump function leading to heart failure. However, Lig3 inactivation did not result in nuclear DNA repair deficiency, indicating essential DNA repair functions of Xrcc1 can occur in the absence of Lig3. Instead, we found that Lig1 was critical for DNA repair, but acted in a cooperative manner with Lig3. Additionally, Lig3 deficiency did not recapitulate the hallmark features of neural Xrcc1 inactivation such as DNA damage-induced cerebellar interneuron loss, further underscoring functional separation of these DNA repair factors. Therefore, our data reveal that the critical biological role of Lig3 is to maintain mtDNA integrity and not Xrcc1-dependent DNA repair.
Figures


References
-
- Ljungquist S, Kenne K, Olsson L, Sandstrom M. Altered DNA ligase III activity in the CHO EM9 mutant. Mutat Res. 1994;314:177–186. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
