

Vascular CD39/ENTPD1 directly promotes tumor cell growth by scavenging extracellular adenosine triphosphate
- PMID: 21390184
- PMCID: PMC3050864
- DOI: 10.1593/neo.101332
Vascular CD39/ENTPD1 directly promotes tumor cell growth by scavenging extracellular adenosine triphosphate
Abstract
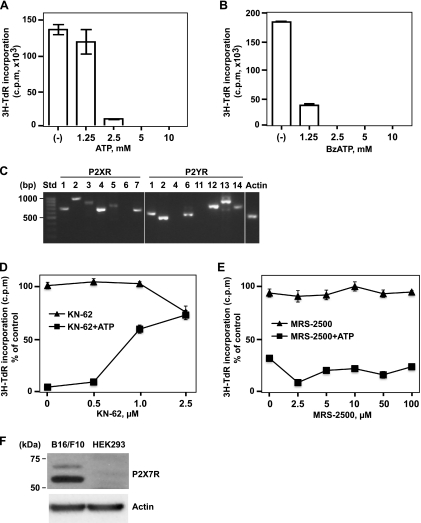
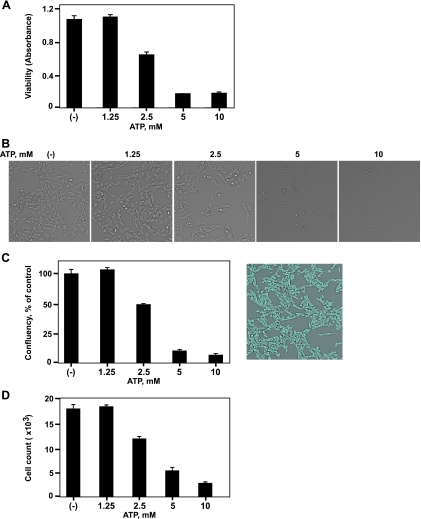
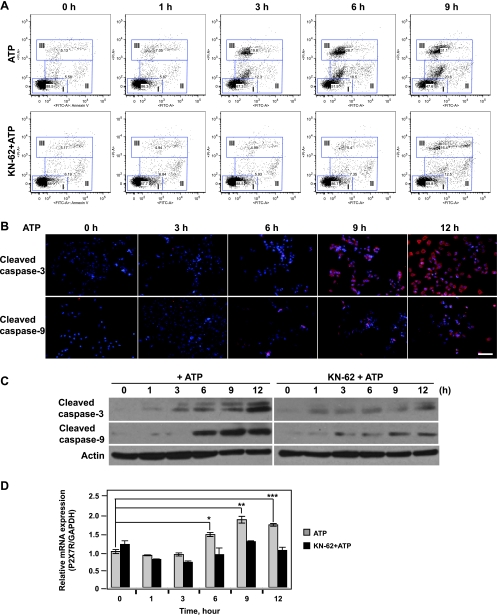
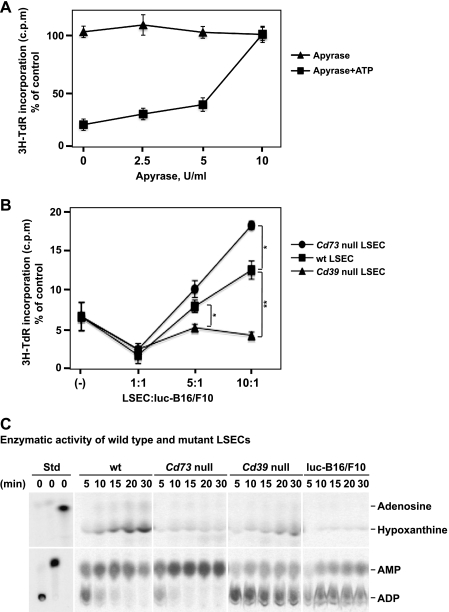
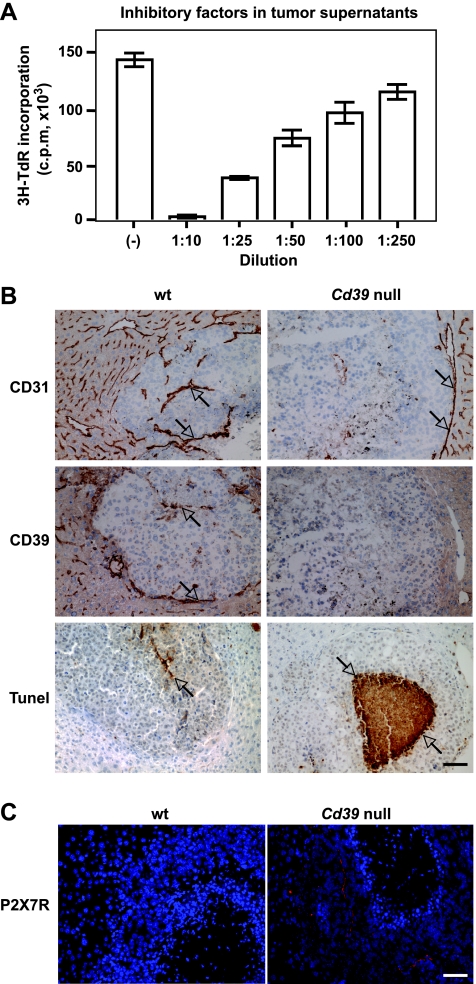
Extracellular adenosine triphosphate (ATP) is known to boost immune responses in the tumor microenvironment but might also contribute directly to cancer cell death. CD39/ENTPD1 is the dominant ectonucleotidase expressed by endothelial cells and regulatory T cells and catalyzes the sequential hydrolysis of ATP to AMP that is further degraded to adenosine by CD73/ecto-5'-nucleotidase. We have previously shown that deletion of Cd39 results in decreased growth of transplanted tumors in mice, as a result of both defective angiogenesis and heightened innate immune responses (secondary to loss of adenosinergic immune suppression). Whether alterations in local extracellular ATP and adenosine levels as a result of CD39 bioactivity directly affect tumor growth and cytotoxicity has not been investigated to date. We show here that extracellular ATP exerts antitumor activity by directly inhibiting cell proliferation and promoting cancer cell death. ATP-induced antiproliferative effects and cell death are, in large part, mediated through P2X(7) receptor signaling. Tumors in Cd39 null mice exhibit increased necrosis in association with P2X(7) expression. We further demonstrate that exogenous soluble NTPDase, or CD39 expression by cocultured liver sinusoidal endothelial cells, stimulates tumor cell proliferation and limits cell death triggered by extracellular ATP. Collectively, our findings indicate that local expression of CD39 directly promotes tumor cell growth by scavenging extracellular ATP. Pharmacological or targeted inhibition of CD39 enzymatic activity may find utility as an adjunct therapy in cancer management.
Figures
References
-
- Burnstock G, Knight GE. Cellular distribution and functions of P2 receptor subtypes in different systems. Int Rev Cytol. 2004;240:31–304. - PubMed
-
- Shabbir M, Burnstock G. Purinergic receptor-mediated effects of adenosine 5′-triphosphate in urological malignant diseases. Int J Urol. 2009;16:143–150. - PubMed
-
- Burnstock G. Purinergic P2 receptors as targets for novel analgesics. Pharmacol Ther. 2006;110:433–454. - PubMed
-
- Burnstock G, Kennedy C. Is there a basis for distinguishing two types of P2-purinoceptor? Gen Pharmacol. 1985;16:433–440. - PubMed
-
- Burnstock G. Purinergic signalling—an overview. Novartis Found Symp. 2006;276:26–48. discussion 48–57, 275–281. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials