

Murine melanoma-infiltrating dendritic cells are defective in antigen presenting function regardless of the presence of CD4CD25 regulatory T cells
- PMID: 21390236
- PMCID: PMC3048402
- DOI: 10.1371/journal.pone.0017515
Murine melanoma-infiltrating dendritic cells are defective in antigen presenting function regardless of the presence of CD4CD25 regulatory T cells
Abstract
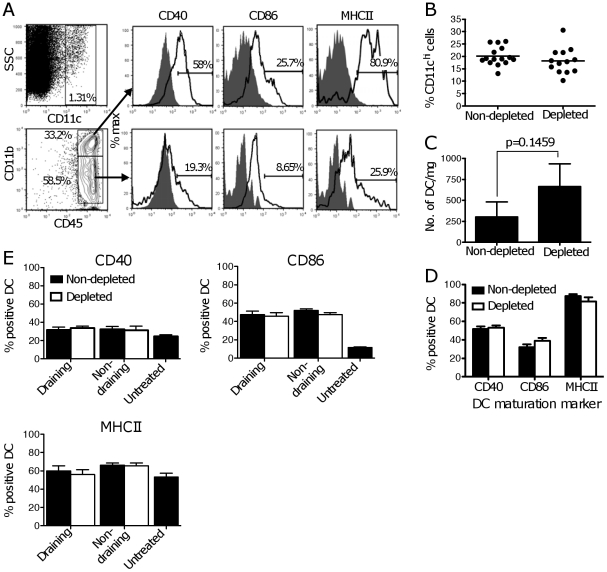
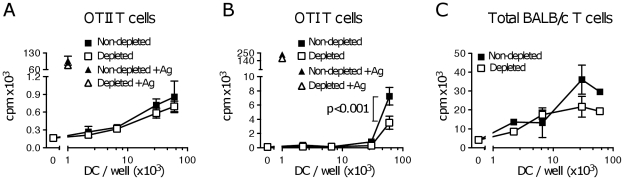
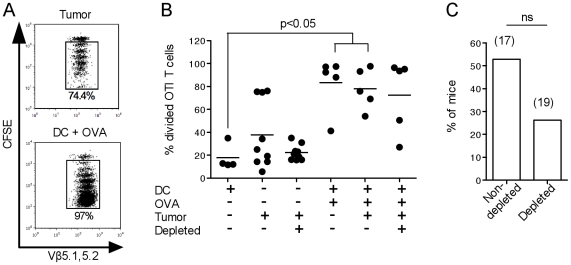
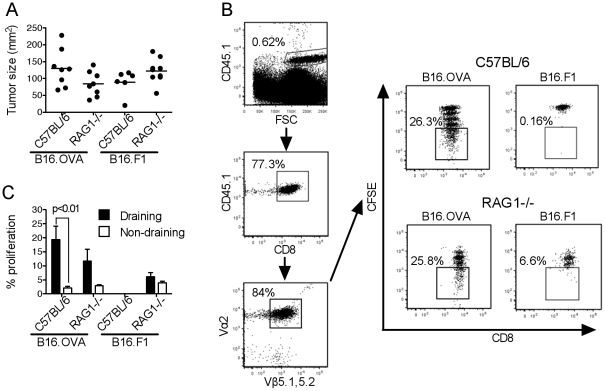
Tumor-infiltrating dendritic cells are often ineffective at presenting tumor-derived antigen in vivo, a defect usually ascribed to the suppressive tumor environment. We investigated the effects of depleting CD4(+)CD25(+) "natural" regulatory T cells (Treg) on the frequency, phenotype and function of total dendritic cell populations in B16.OVA tumors and in tumor-draining lymph nodes. Intraperitoneal injection of the anti-CD25 monoclonal antibody PC61 reduced Treg frequency in blood and tumors, but did not affect the frequency of tumor-infiltrating dendritic cells, or their expression of CD40, CD86 and MHCII. Tumor-infiltrating dendritic cells from PC61-treated or untreated mice induced the proliferation of allogeneic T cells in vitro, but could not induce proliferation of OVA-specific OTI and OTII T cells unless specific peptide antigen was added in culture. Some proliferation of naïve, OVA-specific OTI T cells, but not OTII T cells, was observed in the tumor-draining LN of mice carrying B16.OVA tumors, however, this was not improved by PC61 treatment. Experiments using RAG1(-/-) hosts adoptively transferred with OTI and CD25-depleted OTII cells also failed to show improved OTI and OTII T cell proliferation in vivo compared to C57BL/6 hosts. We conclude that the defective presentation of B16.OVA tumor antigen by tumor-infiltrating dendritic cells and in the tumor-draining lymph node is not due to the presence of "natural" CD4(+)CD25(+) Treg.
Conflict of interest statement
Figures

References
-
- Vicari AP, Caux C, Trinchieri G. Tumour escape from immune surveillance through dendritic cell inactivation. Semin Cancer Biol. 2002;12:33–42. - PubMed
-
- Chiodoni C, Paglia P, Stoppacciaro A, Rodolfo M, Parenza M, et al. Dendritic cells infiltrating tumors cotransduced with granulocyte/macrophage colony-stimulating factor (GM-CSF) and CD40 ligand genes take up and present endogenous tumor-associated antigens, and prime naive mice for a cytotoxic T lymphocyte response. J Exp Med. 1999;190:125–133. - PMC - PubMed
-
- Iwamoto M, Shinohara H, Miyamoto A, Okuzawa M, Mabuchi H, et al. Prognostic value of tumor-infiltrating dendritic cells expressing CD83 in human breast carcinomas. Int J Cancer. 2003;104:92–97. - PubMed
-
- Sandel MH, Dadabayev AR, Menon AG, Morreau H, Melief CJ, et al. Prognostic value of tumor-infiltrating dendritic cells in colorectal cancer: role of maturation status and intratumoral localization. Clin Cancer Res. 2005;11:2576–2582. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
