

Dynamically-driven inactivation of the catalytic machinery of the SARS 3C-like protease by the N214A mutation on the extra domain
- PMID: 21390281
- PMCID: PMC3044768
- DOI: 10.1371/journal.pcbi.1001084
Dynamically-driven inactivation of the catalytic machinery of the SARS 3C-like protease by the N214A mutation on the extra domain
Abstract
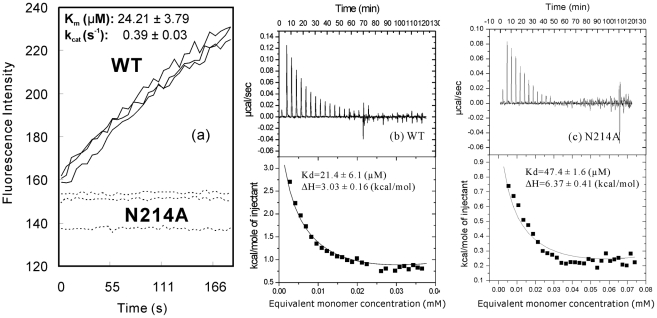
Despite utilizing the same chymotrypsin fold to host the catalytic machinery, coronavirus 3C-like proteases (3CLpro) noticeably differ from picornavirus 3C proteases in acquiring an extra helical domain in evolution. Previously, the extra domain was demonstrated to regulate the catalysis of the SARS-CoV 3CLpro by controlling its dimerization. Here, we studied N214A, another mutant with only a doubled dissociation constant but significantly abolished activity. Unexpectedly, N214A still adopts the dimeric structure almost identical to that of the wild-type (WT) enzyme. Thus, we conducted 30-ns molecular dynamics (MD) simulations for N214A, WT, and R298A which we previously characterized to be a monomer with the collapsed catalytic machinery. Remarkably, three proteases display distinctive dynamical behaviors. While in WT, the catalytic machinery stably retains in the activated state; in R298A it remains largely collapsed in the inactivated state, thus implying that two states are not only structurally very distinguishable but also dynamically well separated. Surprisingly, in N214A the catalytic dyad becomes dynamically unstable and many residues constituting the catalytic machinery jump to sample the conformations highly resembling those of R298A. Therefore, the N214A mutation appears to trigger the dramatic change of the enzyme dynamics in the context of the dimeric form which ultimately inactivates the catalytic machinery. The present MD simulations represent the longest reported so far for the SARS-CoV 3CLpro, unveiling that its catalysis is critically dependent on the dynamics, which can be amazingly modulated by the extra domain. Consequently, mediating the dynamics may offer a potential avenue to inhibit the SARS-CoV 3CLpro.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








References
-
- Anand K, Ziebuhr J, Wadhwani P, Mesters JR, Hilgenfeld R. Coronavirus main proteinase (3CLpro) structure: basis for design of anti- SARS drugs. Science. 2003;300:1763–1767. - PubMed
-
- Allaire M, Chernaia MM, Malcolm BA, James MN. Picornaviral 3C cysteine proteinases have a fold similar to chymotrypsin-like serine proteinases. Nature. 1994;369:72–6. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
