

Ciliary transition zone activation of phosphorylated Tctex-1 controls ciliary resorption, S-phase entry and fate of neural progenitors
- PMID: 21394082
- PMCID: PMC4018803
- DOI: 10.1038/ncb2218
Ciliary transition zone activation of phosphorylated Tctex-1 controls ciliary resorption, S-phase entry and fate of neural progenitors
Abstract
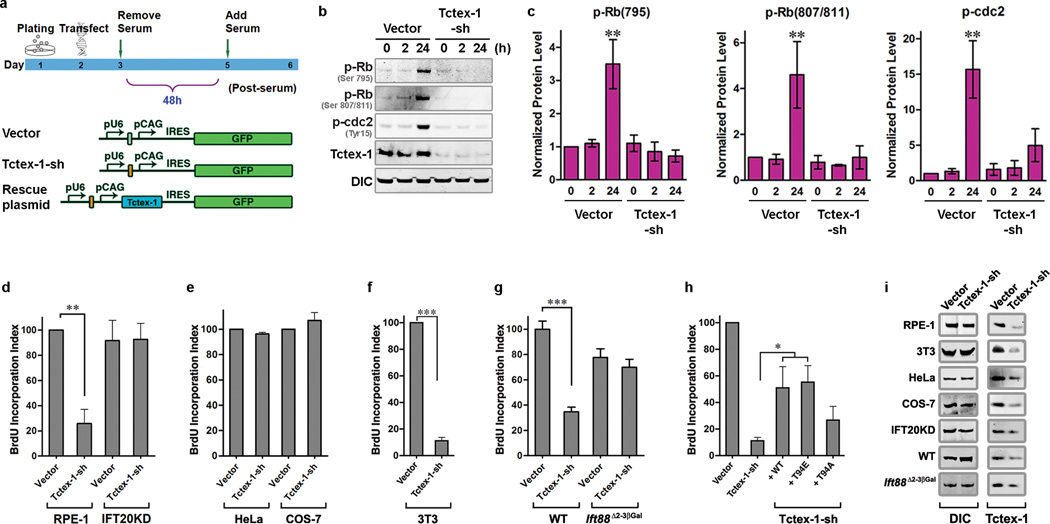
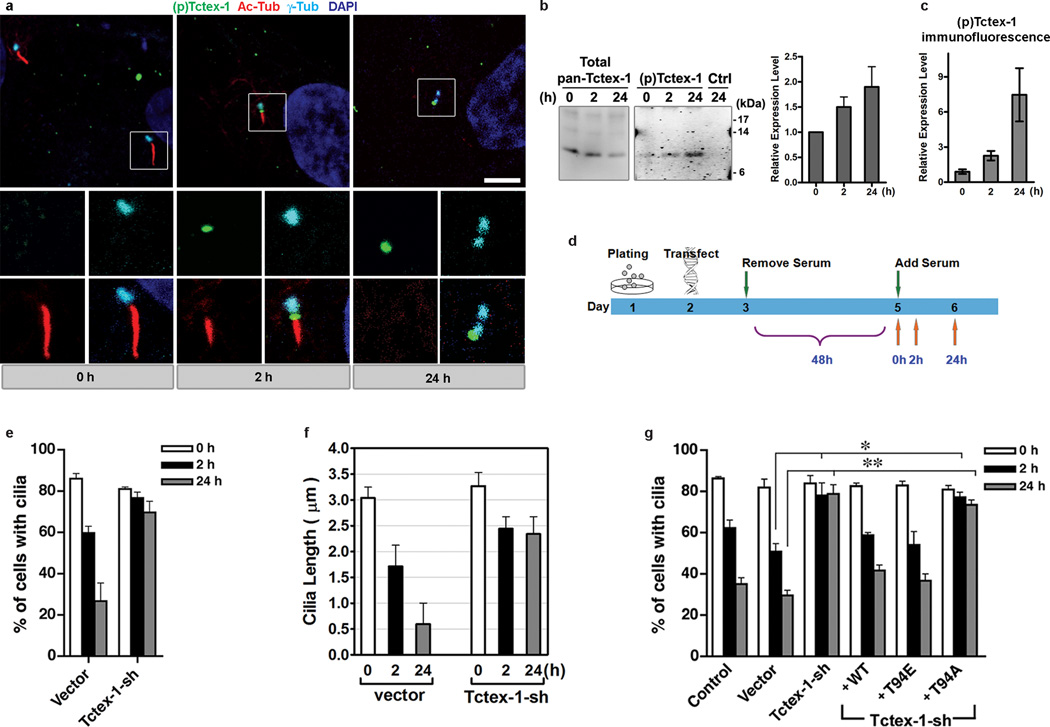
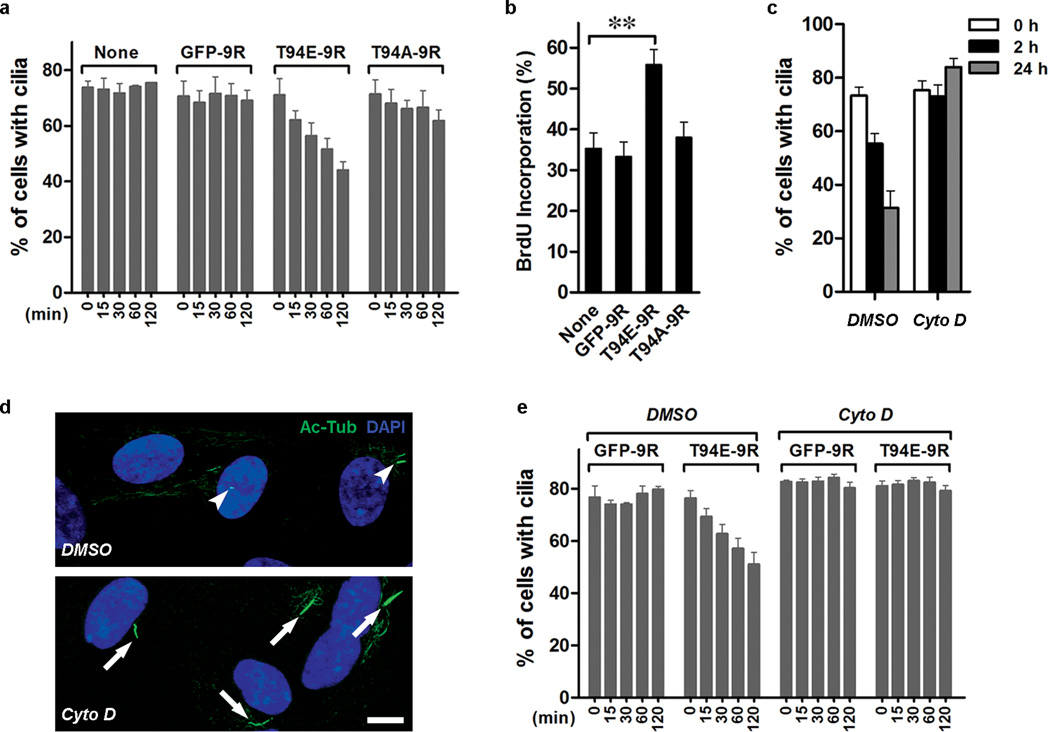
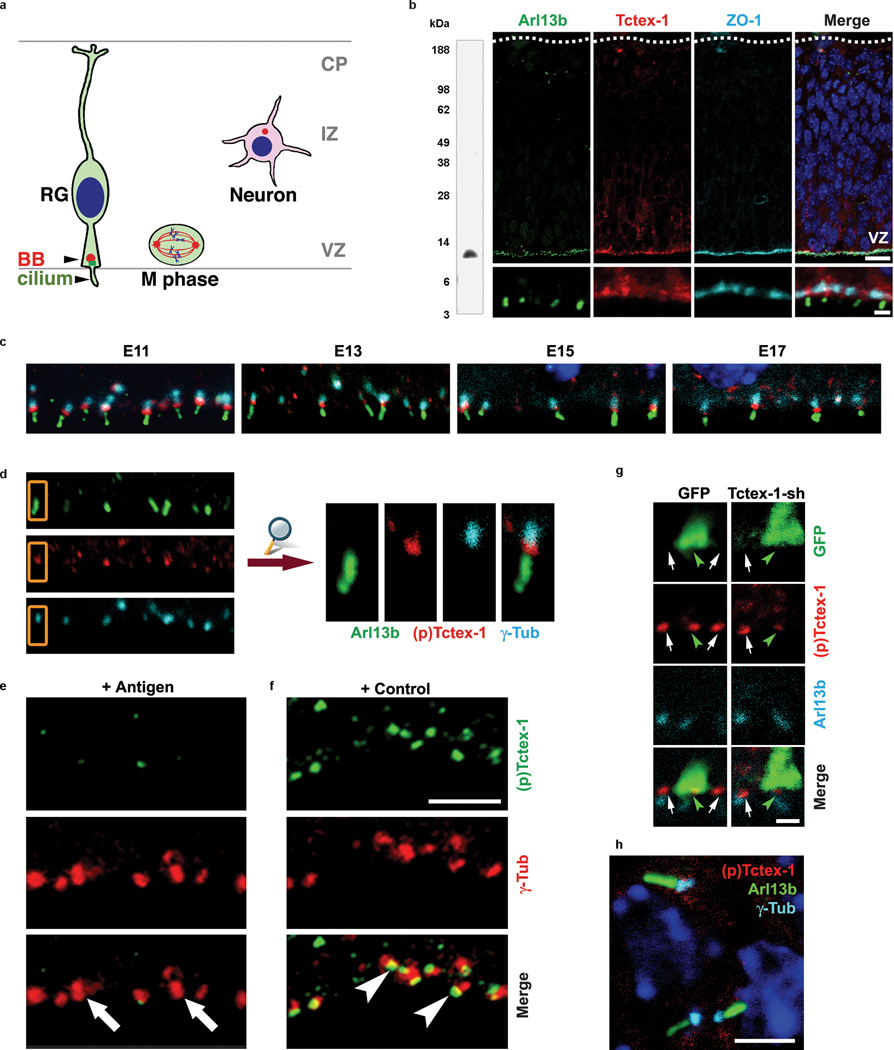
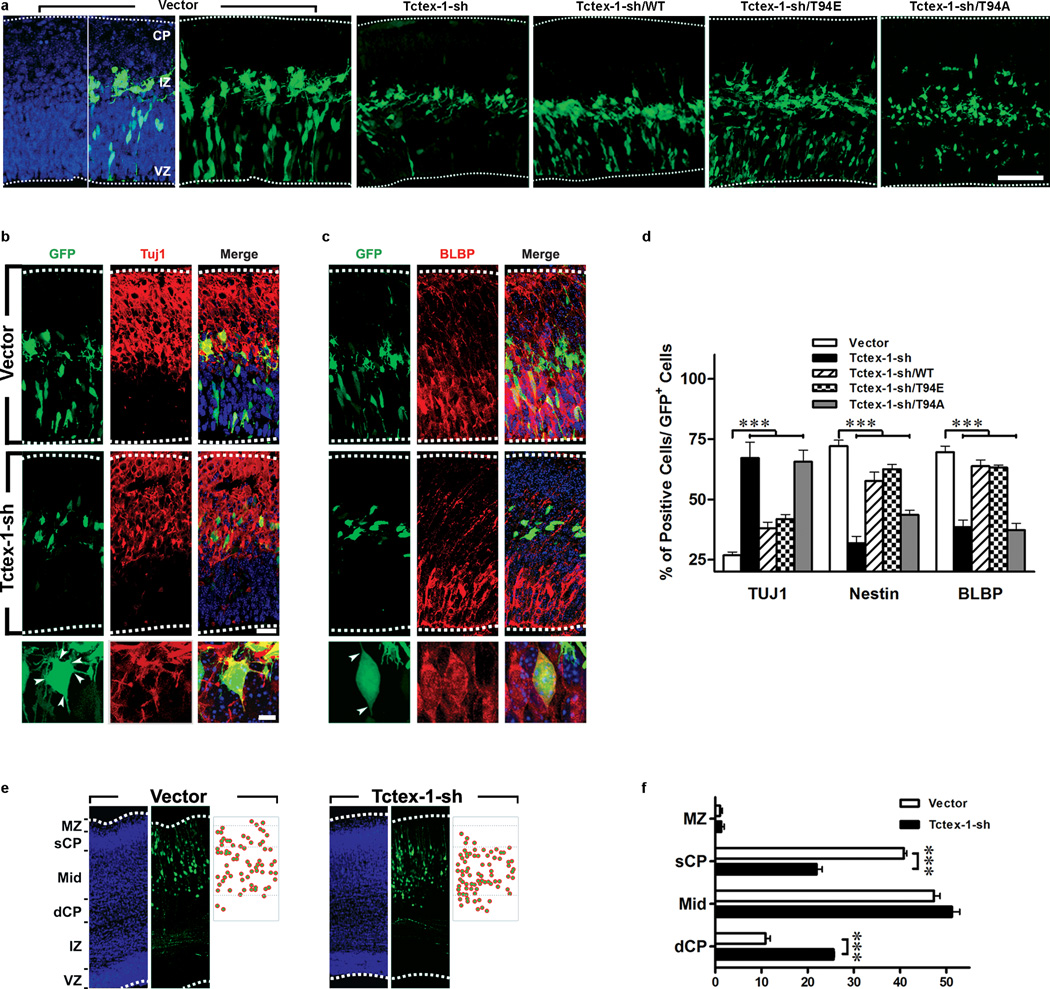
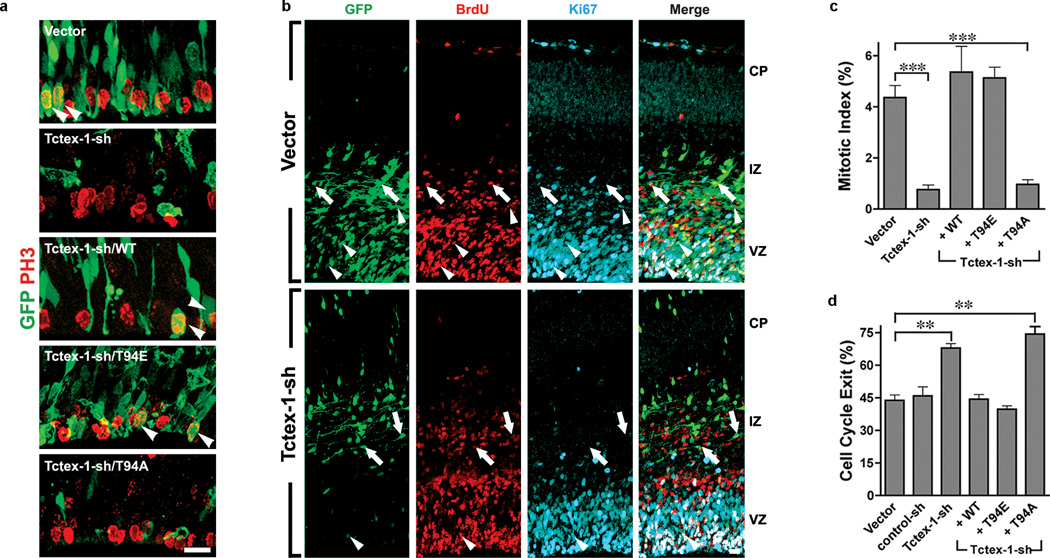
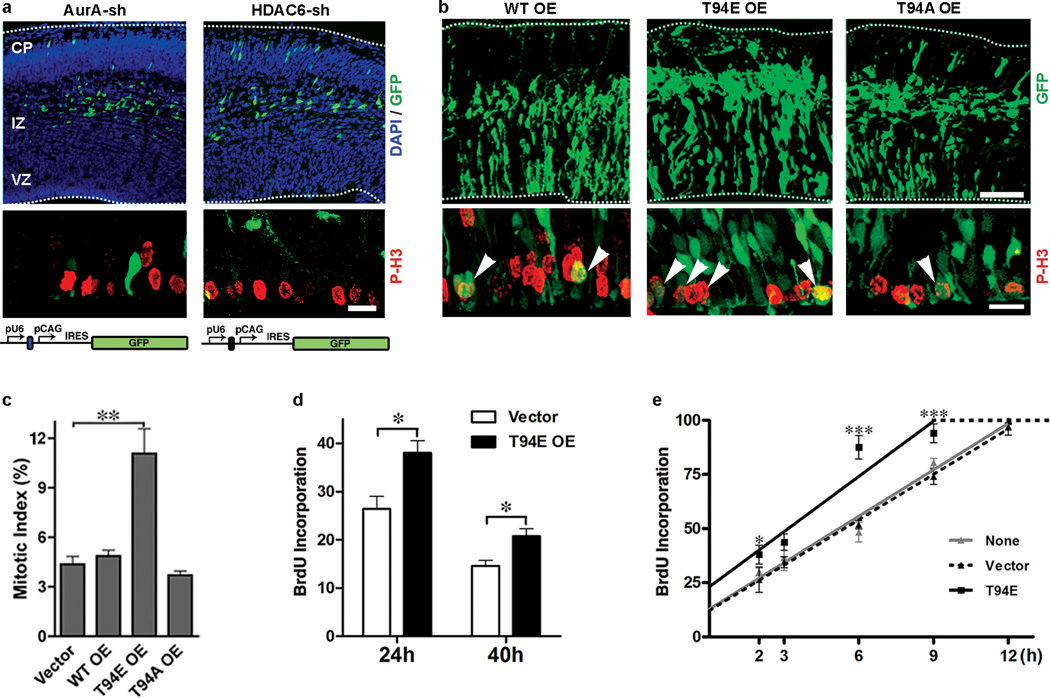
Primary cilia are displayed during the G(0)/G(1) phase of many cell types. Cilia are resorbed as cells prepare to re-enter the cell cycle, but the causal and molecular link between these two cellular events remains unclear. We show that Tctex-1 phosphorylated at Thr 94 is recruited to ciliary transition zones before S-phase entry and has a pivotal role in both ciliary disassembly and cell cycle progression. However, the role of Tctex-1 in S-phase entry is dispensable in non-ciliated cells. Exogenously adding a phospho-mimic Tctex-1(T94E) mutant accelerates cilium disassembly and S-phase entry. These results support a model in which the cilia act as a brake to prevent cell cycle progression. Mechanistic studies show the involvement of actin dynamics in Tctex-1-regulated cilium resorption. Tctex-1 phosphorylated at Thr 94 is also selectively enriched at the ciliary transition zones of cortical neural progenitors, and has a key role in controlling G(1) length, cell cycle entry and fate determination of these cells during corticogenesis.
© 2011 Macmillan Publishers Limited. All rights reserved
Figures
Comment in
-
Do cilia put brakes on the cell cycle?Nat Cell Biol. 2011 Apr;13(4):340-2. doi: 10.1038/ncb0411-340. Nat Cell Biol. 2011. PMID: 21460803
References
-
- Pan J, Snell W. The primary cilium: keeper of the key to cell division. Cell. 2007;129:1255–1257. - PubMed
-
- Alvarez-Buylla A, Garcia-Verdugo JM, Tramontin AD. A unified hypothesis on the lineage of neural stem cells. Nat Rev Neurosci. 2001;2:287–293. - PubMed
-
- Lange C, Huttner WB, Calegari F. Cdk4/cyclinD1 overexpression in neural stem cells shortens G1, delays neurogenesis, and promotes the generation and expansion of basal progenitors. Cell Stem Cell. 2009;5:320–331. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
