

Blood vessel tubulogenesis requires Rasip1 regulation of GTPase signaling
- PMID: 21396893
- PMCID: PMC3078994
- DOI: 10.1016/j.devcel.2011.02.010
Blood vessel tubulogenesis requires Rasip1 regulation of GTPase signaling
Abstract
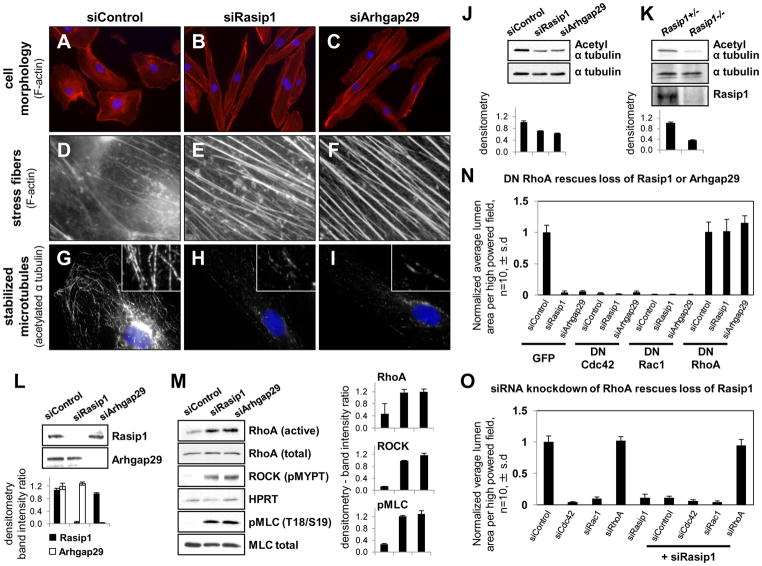
Cardiovascular function depends on patent blood vessel formation by endothelial cells (ECs). However, the mechanisms underlying vascular "tubulogenesis" are only beginning to be unraveled. We show that endothelial tubulogenesis requires the Ras interacting protein 1, Rasip1, and its binding partner, the RhoGAP Arhgap29. Mice lacking Rasip1 fail to form patent lumens in all blood vessels, including the early endocardial tube. Rasipl null angioblasts fail to properly localize the polarity determinant Par3 and display defective cell polarity, resulting in mislocalized junctional complexes and loss of adhesion to extracellular matrix (ECM). Similarly, depletion of either Rasip1 or Arhgap29 in cultured ECs blocks in vitro lumen formation, fundamentally alters the cytoskeleton, and reduces integrin-dependent adhesion to ECM. These defects result from increased RhoA/ROCK/myosin II activity and blockade of Cdc42 and Rac1 signaling. This study identifies Rasip1 as a unique, endothelial-specific regulator of Rho GTPase signaling, which is essential for blood vessel morphogenesis.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures






Comment in
-
LUMENating blood vessels.Dev Cell. 2011 Apr 19;20(4):412-4. doi: 10.1016/j.devcel.2011.03.020. Dev Cell. 2011. PMID: 21497753 Free PMC article.
References
-
- Aird WC. Phenotypic heterogeneity of the endothelium: II. Representative vascular beds. Circ Res. 2007;100:174–190. - PubMed
-
- Bayless KJ, Davis GE. The Cdc42 and Rac1 GTPases are required for capillary lumen formation in three-dimensional extracellular matrices. J Cell Sci. 2002;115:1123–1136. - PubMed
-
- Bayless KJ, Davis GE. Microtubule depolymerization rapidly collapses capillary tube networks in vitro and angiogenic vessels in vivo through the small GTPase Rho. J Biol Chem. 2004;279:11686–11695. - PubMed
-
- Bazzoni G, Shih DT, Buck CA, Hemler ME. Monoclonal antibody 9EG7 defines a novel beta 1 integrin epitope induced by soluble ligand and manganese, but inhibited by calcium. J Biol Chem. 1995;270:25570–25577. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
