

Nuclear localization of pyruvate dehydrogenase complex-E2 (PDC-E2), a mitochondrial enzyme, and its role in signal transducer and activator of transcription 5 (STAT5)-dependent gene transcription
- PMID: 21397011
- PMCID: PMC3096605
- DOI: 10.1016/j.cellsig.2011.03.004
Nuclear localization of pyruvate dehydrogenase complex-E2 (PDC-E2), a mitochondrial enzyme, and its role in signal transducer and activator of transcription 5 (STAT5)-dependent gene transcription
Abstract
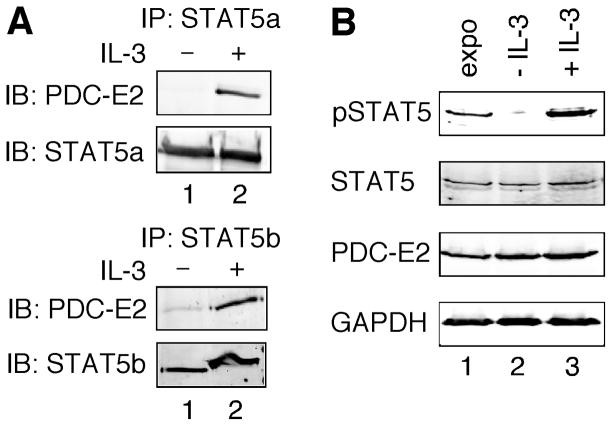
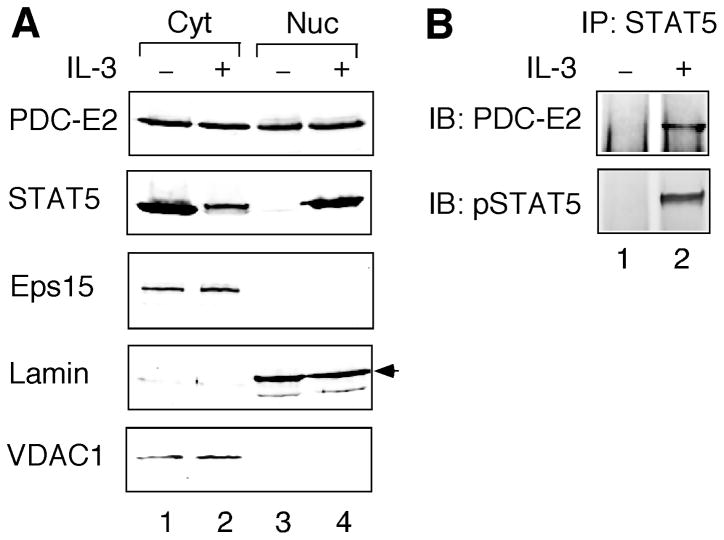
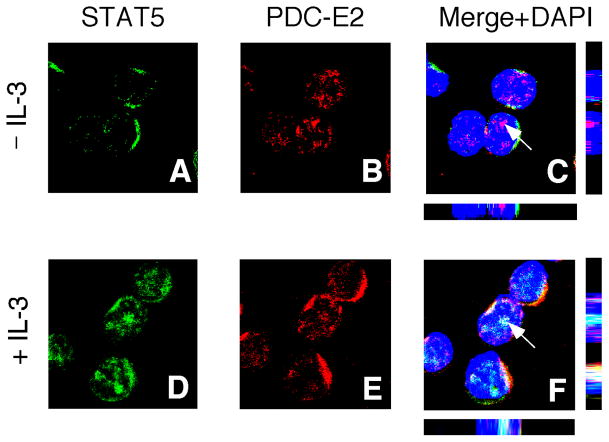
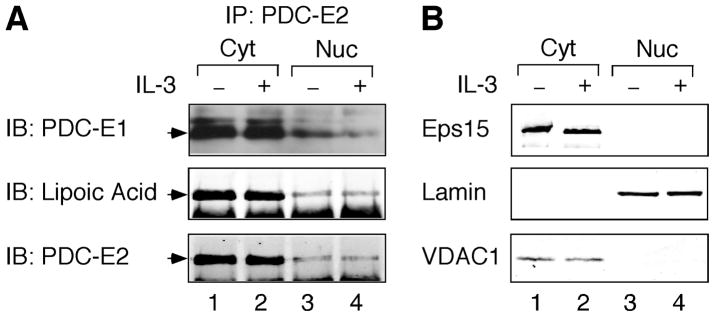
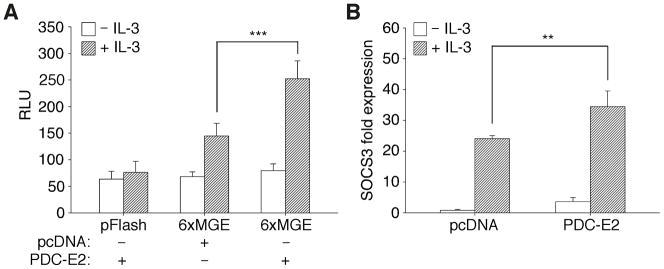
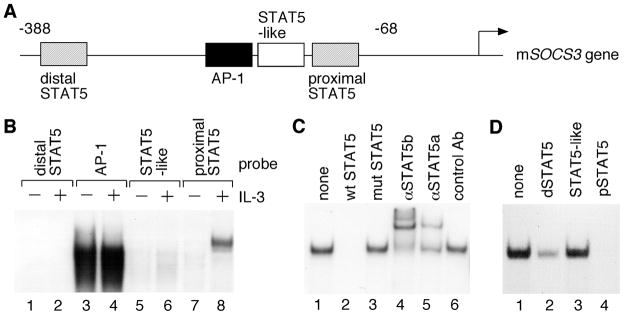
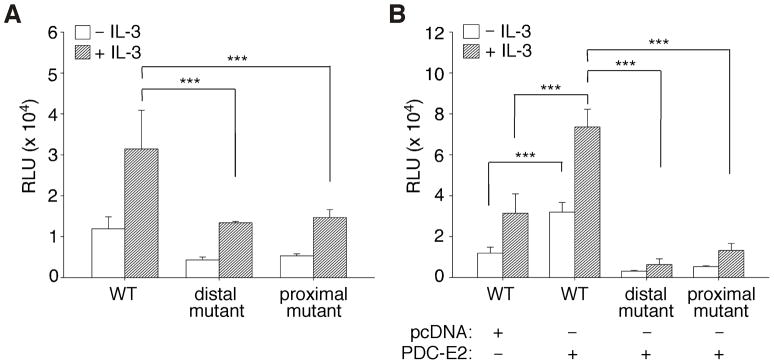
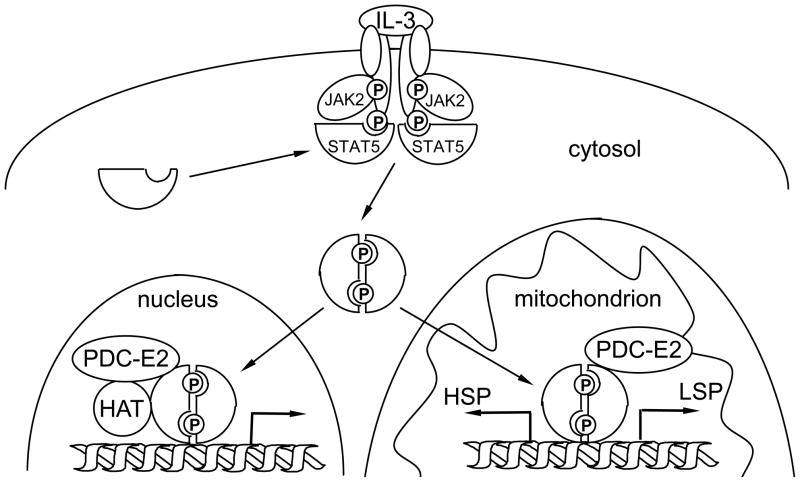
STAT (signal transducer and activator of transcription) proteins play a critical role in cellular response to a wide variety of cytokines and growth factors by regulating specific nuclear genes. STAT-dependent gene transcription can be finely tuned through the association with co-factors in the nucleus. We showed previously that STAT5 (including 5a and 5b) specifically interacts with a mitochondrial enzyme PDC-E2 (E2 subunit of pyruvate dehydrogenase complex) in both leukemic T cells and cytokine-stimulated cells. However, the functional significance of this novel association remains largely unknown. Here we report that PDC-E2 may function as a co-activator in STAT5-dependent nuclear gene expression. Subcellular fractionation analysis revealed that a substantial amount of PDC-E2 was constitutively present in the nucleus of BaF3, an interleukin-3 (IL-3)-dependent cell line. IL-3-induced tyrosine-phosphorylated STAT5 associated with nuclear PDC-E2 in co-immunoprecipitation analysis. These findings were confirmed by confocal immunofluorescence microscopy showing constant nuclear localization of PDC-E2 and its co-localization with STAT5 after IL-3 stimulation. Similar to mitochondrial PDC-E2, nuclear PDC-E2 was lipoylated and associated with PDC-E1. Overexpression of PDC-E2 in BaF3 cells augmented IL-3-induced STAT5 activity as measured by reporter assay with consensus STAT5-binding sites. Consistent with the reporter data, PDC-E2 overexpression in BaF3 cells led to elevated mRNA levels of endogenous SOCS3 (suppressor of cytokine signaling 3) gene, a known STAT5 target. We further identified two functional STAT5-binding sites in the SOCS3 gene promoter important for its IL-3-inducibility. The observation that both cis-acting elements were essential to detect the stimulatory effect by PDC-E2 strongly supports the role of PDC-E2 in up-regulating the transactivating ability of STAT5. All together, our results reveal a novel function of PDC-E2 in the nucleus. It also raises the possibility of nuclear-mitochondrial crosstalk through the interaction between STAT5 and PDC-E2.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Levy DE, Darnell JE., Jr Nat Rev Mol Cell Biol. 2002;3:651–662. - PubMed
-
- Schindler C, Levy DE, Decker T. J Biol Chem. 2007;282:20059–20063. - PubMed
-
- Lanning NJ, Carter-Su C. Rev Endocr Metab Dis. 2006;7:225–235. - PubMed
-
- Yu H, Jove R. Nat Rev Cancer. 2004;4:97–105. - PubMed
-
- Shuai K. Oncogene. 2000;19:2638–2644. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
