

MITOPLD is a mitochondrial protein essential for nuage formation and piRNA biogenesis in the mouse germline
- PMID: 21397847
- PMCID: PMC3062204
- DOI: 10.1016/j.devcel.2011.01.005
MITOPLD is a mitochondrial protein essential for nuage formation and piRNA biogenesis in the mouse germline
Abstract
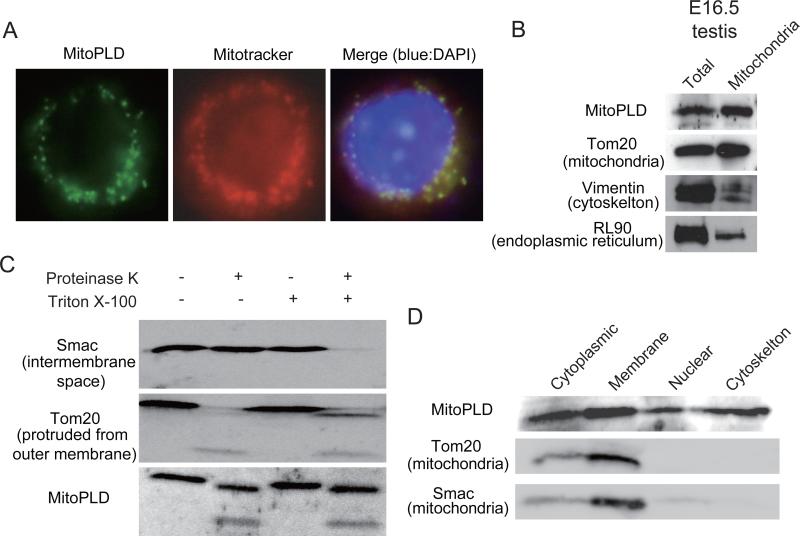
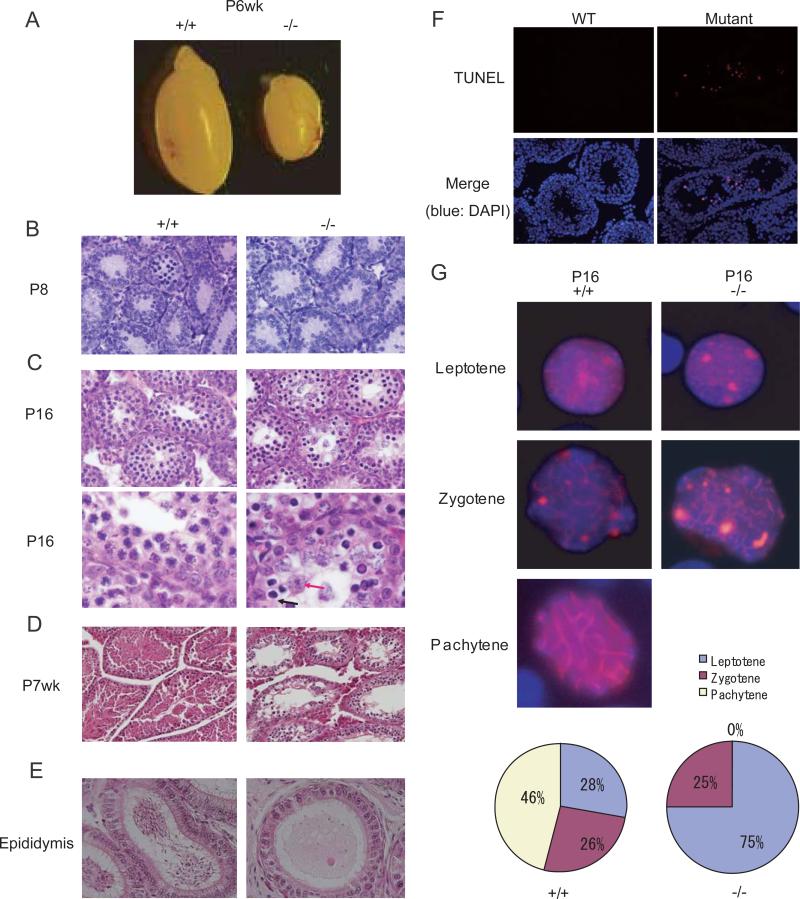
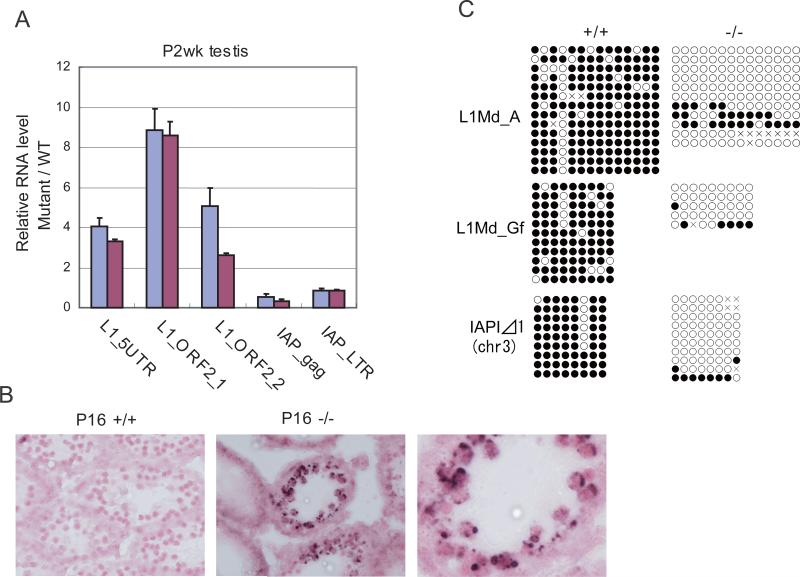
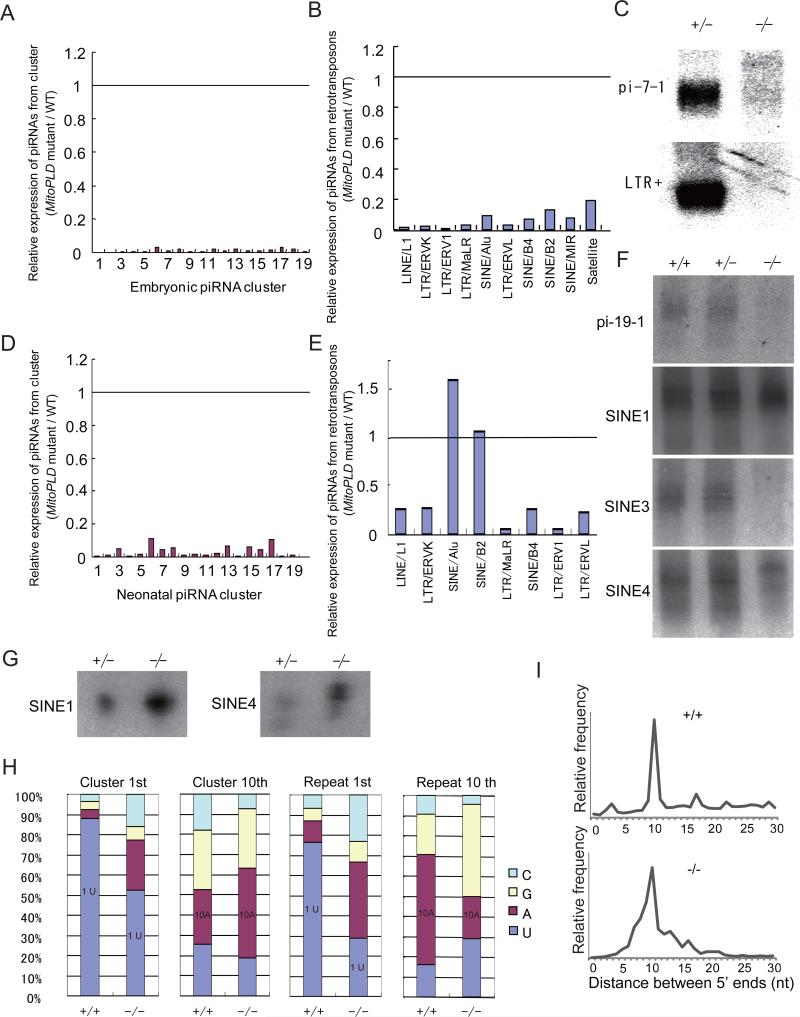
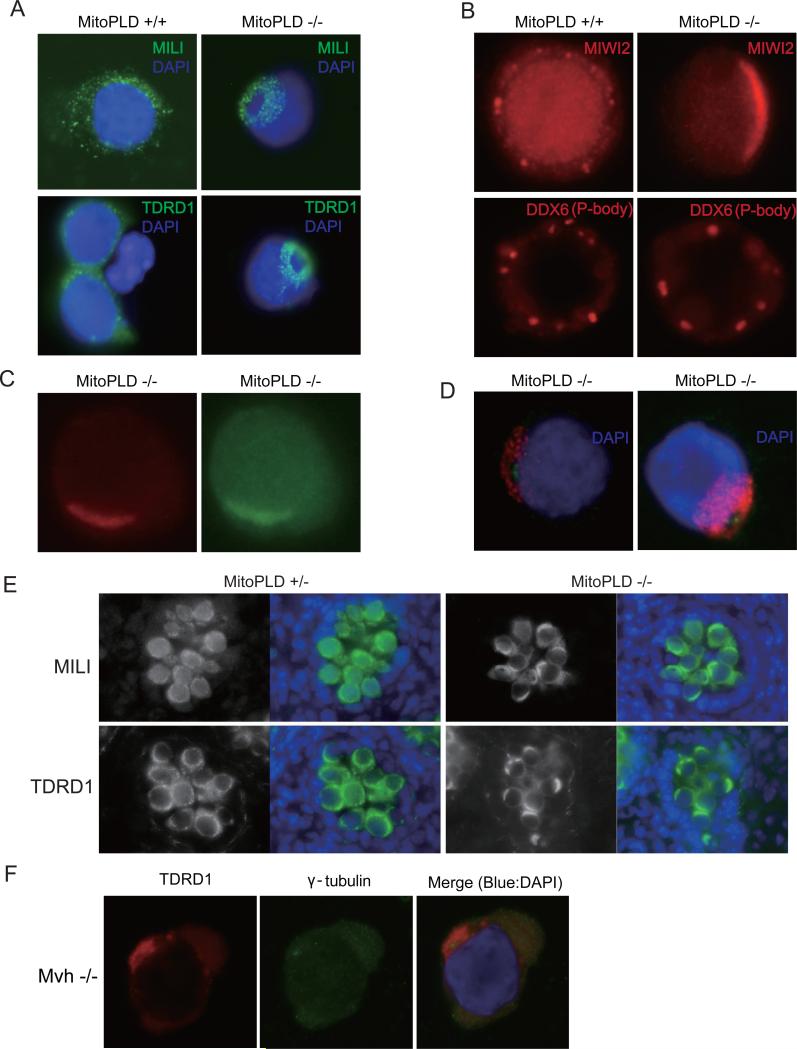
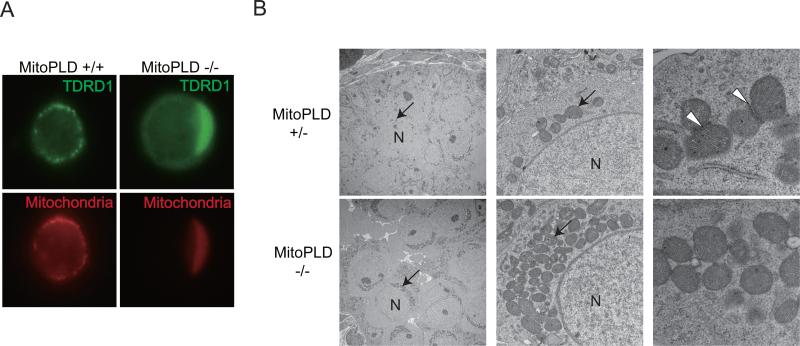
MITOPLD is a member of the phospholipase D superfamily proteins conserved among diverse species. Zucchini (Zuc), the Drosophila homolog of MITOPLD, has been implicated in primary biogenesis of Piwi-interacting RNAs (piRNAs). By contrast, MITOPLD has been shown to hydrolyze cardiolipin in the outer membrane of mitochondria to generate phosphatidic acid, which is a signaling molecule. To assess whether the mammalian MITOPLD is involved in piRNA biogenesis, we generated Mitopld mutant mice. The mice display meiotic arrest during spermatogenesis, demethylation and derepression of retrotransposons, and defects in primary piRNA biogenesis. Furthermore, in mutant germ cells, mitochondria and the components of the nuage, a perinuclear structure involved in piRNA biogenesis/function, are mislocalized to regions around the centrosome, suggesting that MITOPLD may be involved in microtubule-dependent localization of mitochondria and these proteins. Our results indicate a conserved role for MITOPLD/Zuc in the piRNA pathway and link mitochondrial membrane metabolism/signaling to small RNA biogenesis.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Anderson R, Schaible K, Heasman J, Wylie C. Expression of the homophilic adhesion molecule, Ep-CAM, in the mammalian germ line. J Reprod Fertil. 1999;116:379–384. - PubMed
-
- Aravin A, Gaidatzis D, Pfeffer S, Lagos-Quintana M, Landgraf P, Iovino N, Morris P, Brownstein MJ, Kuramochi-Miyagawa S, Nakano T, et al. A novel class of small RNAs bind to MILI protein in mouse testes. Nature. 2006;442:203–207. - PubMed
-
- Aravin AA, Hannon GJ, Brennecke J. The Piwi-piRNA pathway provides an adaptive defense in the transposon arms race. Science. 2007a;318:761–764. - PubMed
-
- Aravin AA, Sachidanandam R, Girard A, Fejes-Toth K, Hannon GJ. Developmentally regulated piRNA clusters implicate MILI in transposon control. Science. 2007b;316:744–747. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
