

Driving forces enable high-titer anaerobic 1-butanol synthesis in Escherichia coli
- PMID: 21398484
- PMCID: PMC3126405
- DOI: 10.1128/AEM.03034-10
Driving forces enable high-titer anaerobic 1-butanol synthesis in Escherichia coli
Abstract
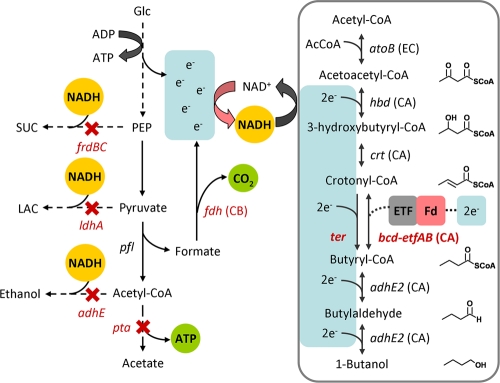
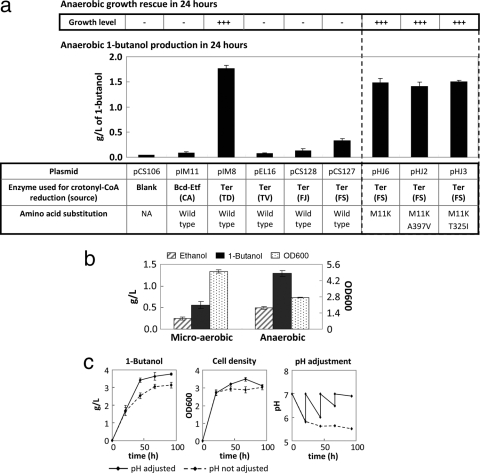
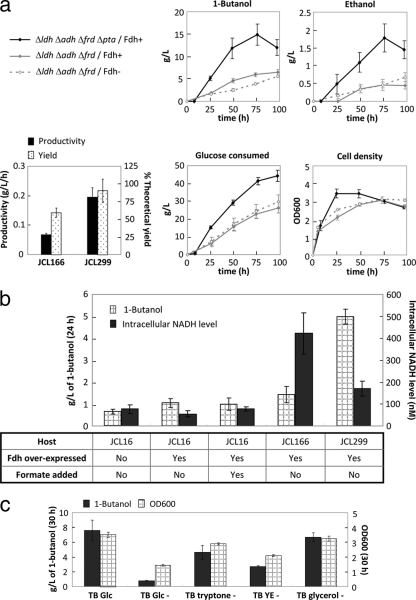
1-Butanol, an important chemical feedstock and advanced biofuel, is produced by Clostridium species. Various efforts have been made to transfer the clostridial 1-butanol pathway into other microorganisms. However, in contrast to similar compounds, only limited titers of 1-butanol were attained. In this work, we constructed a modified clostridial 1-butanol pathway in Escherichia coli to provide an irreversible reaction catalyzed by trans-enoyl-coenzyme A (CoA) reductase (Ter) and created NADH and acetyl-CoA driving forces to direct the flux. We achieved high-titer (30 g/liter) and high-yield (70 to 88% of the theoretical) production of 1-butanol anaerobically, comparable to or exceeding the levels demonstrated by native producers. Without the NADH and acetyl-CoA driving forces, the Ter reaction alone only achieved about 1/10 the level of production. The engineered host platform also enables the selection of essential enzymes with better catalytic efficiency or expression by anaerobic growth rescue. These results demonstrate the importance of driving forces in the efficient production of nonnative products.
Figures


References
-
- Andreadeli A., Platis D., Tishkov V., Popov V., Labrou N. E. 2008. Structure-guided alteration of coenzyme specificity of formate dehydrogenase by saturation mutagenesis to enable efficient utilization of NADP+. FEBS J. 275:3859–3869 - PubMed
-
- Atsumi S., et al. 2008. Metabolic engineering of Escherichia coli for 1-butanol production. Metab. Eng. 10:305–311 - PubMed
-
- Atsumi S., Hanai T., Liao J. C. 2008. Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 451:86–89 - PubMed
-
- Baez A., Cho K. M., Liao J. C. 10 March 2011. High-titer isobutanol production using engineered Escherichia coli: a bioreactor study with in situ product removal. Appl. Microbiol. Biotechnol. doi:10.1007/s00253-011-3173-y - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
