

The MYB96 transcription factor regulates cuticular wax biosynthesis under drought conditions in Arabidopsis
- PMID: 21398568
- PMCID: PMC3082259
- DOI: 10.1105/tpc.111.083485
The MYB96 transcription factor regulates cuticular wax biosynthesis under drought conditions in Arabidopsis
Abstract
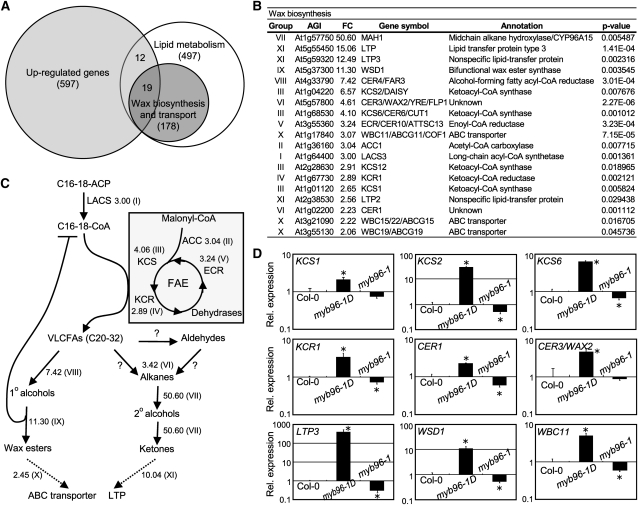
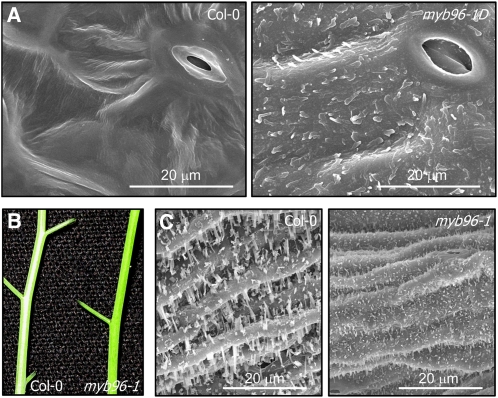
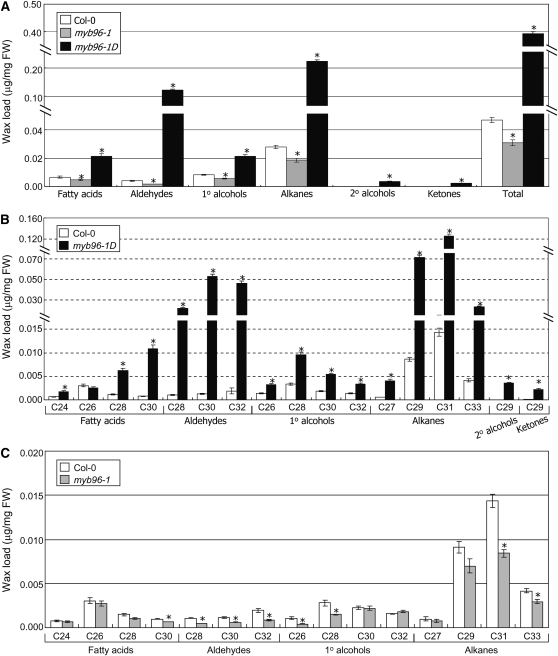
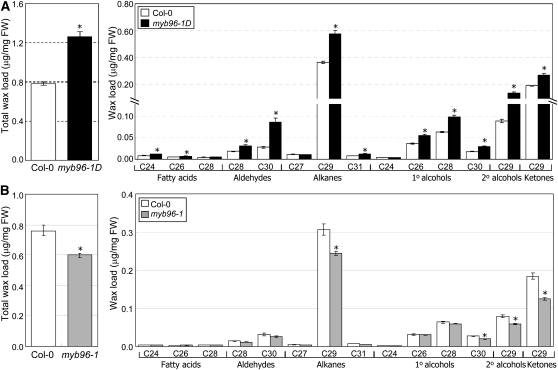
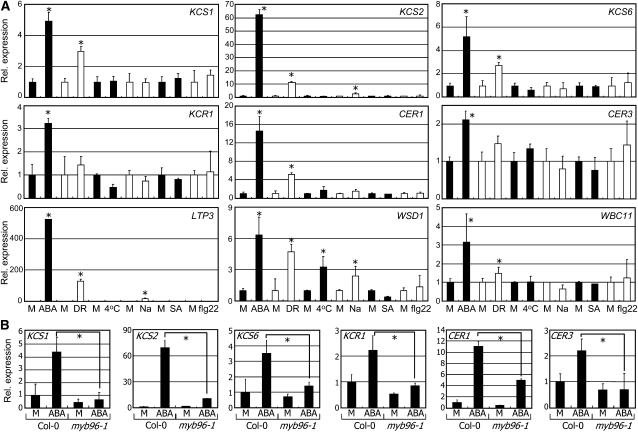
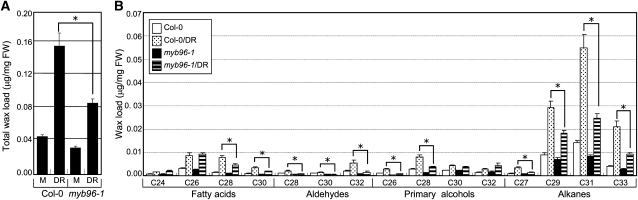
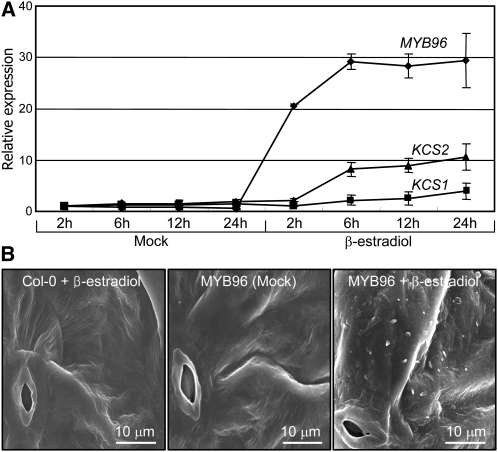
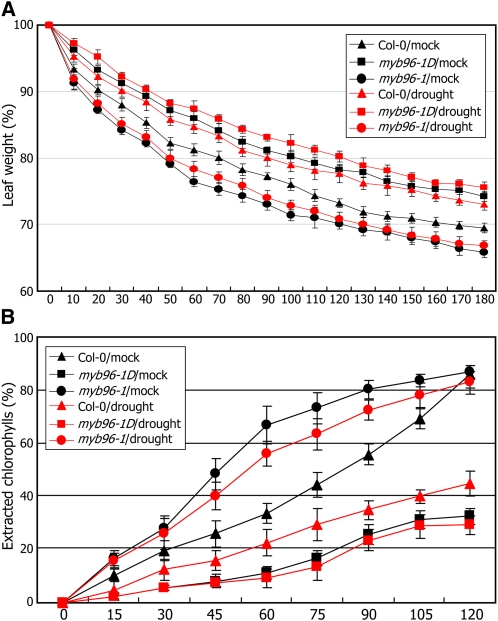
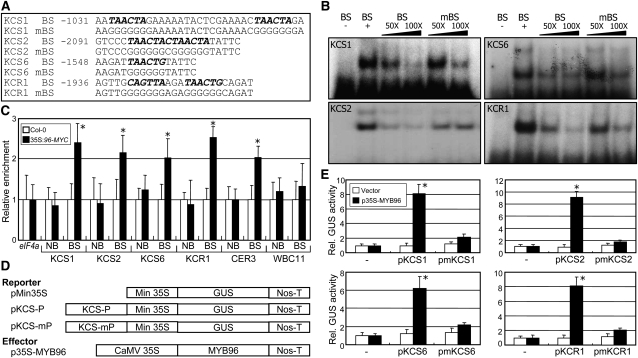
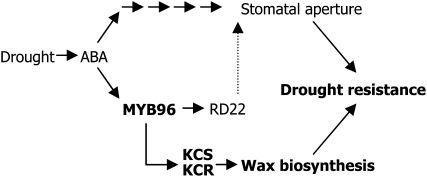
Drought stress activates several defense responses in plants, such as stomatal closure, maintenance of root water uptake, and synthesis of osmoprotectants. Accumulating evidence suggests that deposition of cuticular waxes is also associated with plant responses to cellular dehydration. Yet, how cuticular wax biosynthesis is regulated in response to drought is unknown. We have recently reported that an Arabidopsis thaliana abscisic acid (ABA)-responsive R2R3-type MYB transcription factor, MYB96, promotes drought resistance. Here, we show that transcriptional activation of cuticular wax biosynthesis by MYB96 contributes to drought resistance. Microarray assays showed that a group of wax biosynthetic genes is upregulated in the activation-tagged myb96-1D mutant but downregulated in the MYB96-deficient myb96-1 mutant. Cuticular wax accumulation was altered accordingly in the mutants. In addition, activation of cuticular wax biosynthesis by drought and ABA requires MYB96. By contrast, biosynthesis of cutin monomers was only marginally affected in the mutants. Notably, the MYB96 protein acts as a transcriptional activator of genes encoding very-long-chain fatty acid-condensing enzymes involved in cuticular wax biosynthesis by directly binding to conserved sequence motifs present in the gene promoters. These results demonstrate that ABA-mediated MYB96 activation of cuticular wax biosynthesis serves as a drought resistance mechanism.
Figures
References
-
- Baker C.J., McCormick S.L., Bateman D.F. (1982). Effects of purified cutin esterase upon the permeability and mechanical strength of cutin membranes. Phytopathology 72: 420–423
-
- Barthlott W., Neinhuis C. (1997). Purity of the sacred lotus, or escape from contamination in biological surfaces. Planta 202: 1–8
