

Structural and functional analysis of a plant resistance protein TIR domain reveals interfaces for self-association, signaling, and autoregulation
- PMID: 21402359
- PMCID: PMC3142617
- DOI: 10.1016/j.chom.2011.02.009
Structural and functional analysis of a plant resistance protein TIR domain reveals interfaces for self-association, signaling, and autoregulation
Abstract
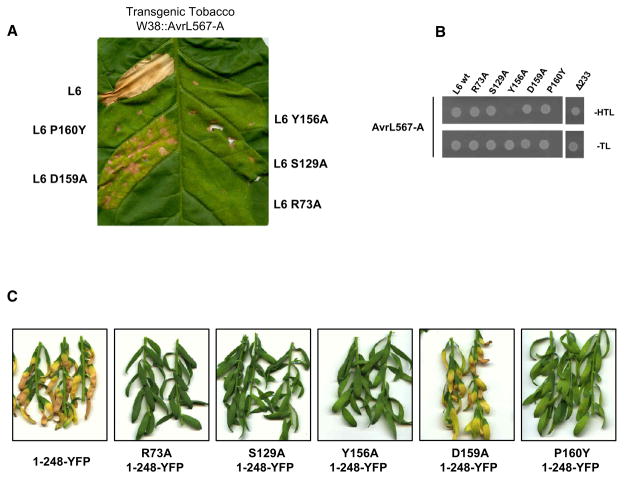
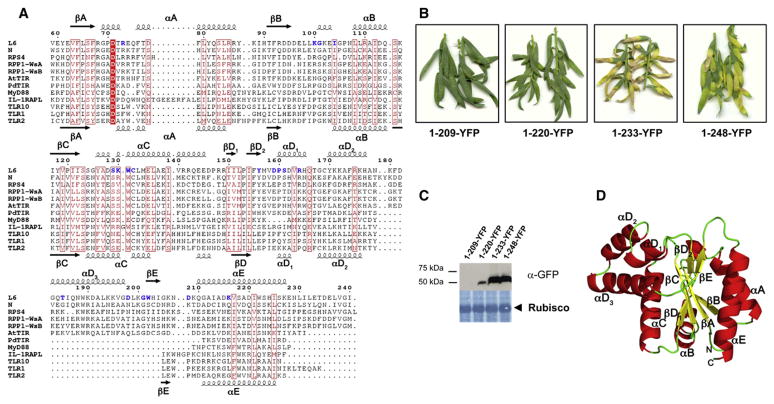
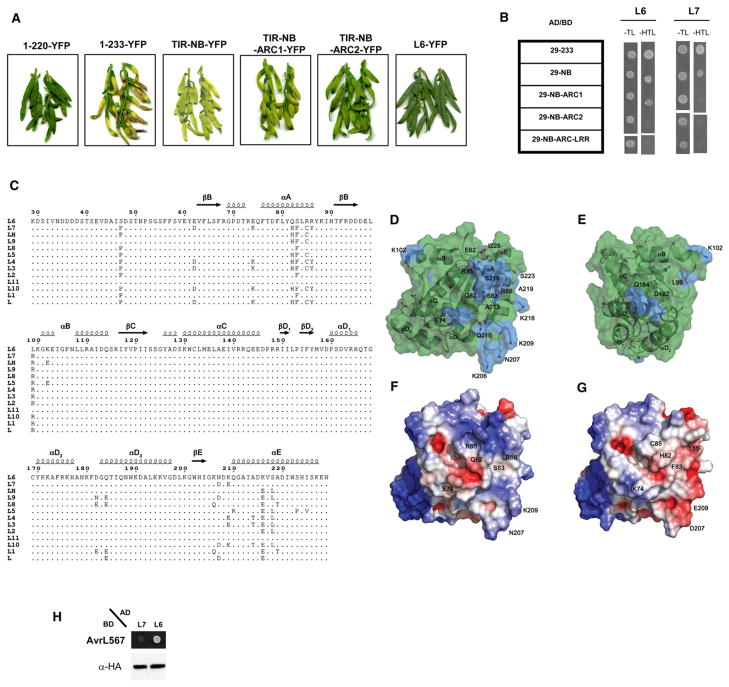
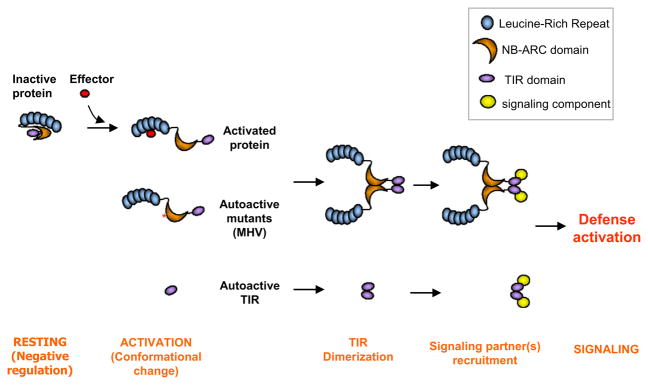
The Toll/interleukin-1 receptor (TIR) domain occurs in animal and plant immune receptors. In the animal Toll-like receptors, homodimerization of the intracellular TIR domain is required for initiation of signaling cascades leading to innate immunity. By contrast, the role of the TIR domain in cytoplasmic nucleotide-binding/leucine-rich repeat (NB-LRR) plant immune resistance proteins is poorly understood. L6 is a TIR-NB-LRR resistance protein from flax (Linum usitatissimum) that confers resistance to the flax rust phytopathogenic fungus (Melampsora lini). We determine the crystal structure of the L6 TIR domain and show that, although dispensable for pathogenic effector protein recognition, the TIR domain alone is both necessary and sufficient for L6 immune signaling. We demonstrate that the L6 TIR domain self-associates, most likely forming a homodimer. Analysis of the structure combined with site-directed mutagenesis suggests that self-association is a requirement for immune signaling and reveals distinct surface regions involved in self-association, signaling, and autoregulation.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures


References
-
- Bao Q, Shi Y. Apoptosome: a platform for the activation of initiator caspases. Cell Death Differ. 2007;14:56–65. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
