

Understanding the role of growth factors in embryonic development: insights from the lens
- PMID: 21402581
- PMCID: PMC3061110
- DOI: 10.1098/rstb.2010.0339
Understanding the role of growth factors in embryonic development: insights from the lens
Abstract
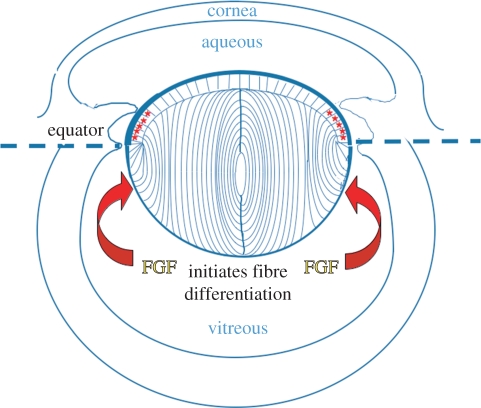
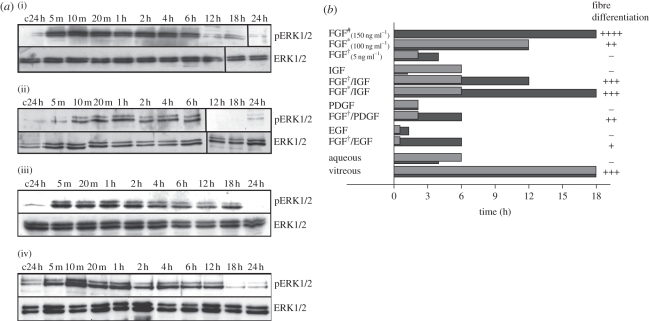
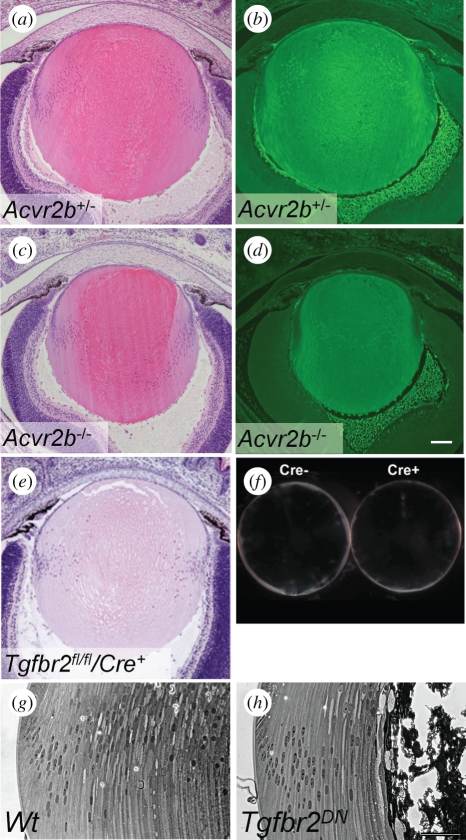
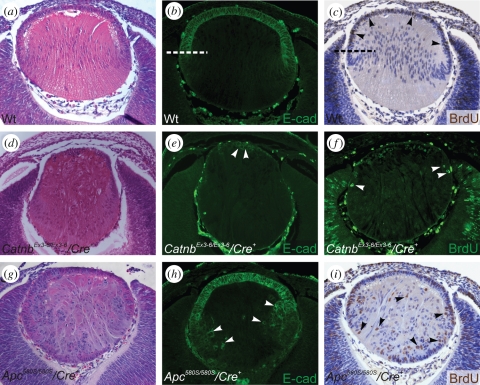
Growth factors play key roles in influencing cell fate and behaviour during development. The epithelial cells and fibre cells that arise from the lens vesicle during lens morphogenesis are bathed by aqueous and vitreous, respectively. Vitreous has been shown to generate a high level of fibroblast growth factor (FGF) signalling that is required for secondary lens fibre differentiation. However, studies also show that FGF signalling is not sufficient and roles have been identified for transforming growth factor-β and Wnt/Frizzled families in regulating aspects of fibre differentiation. In the case of the epithelium, key roles for Wnt/β-catenin and Notch signalling have been demonstrated in embryonic development, but it is not known if other factors are required for its formation and maintenance. This review provides an overview of current knowledge about growth factor regulation of differentiation and maintenance of lens cells. It also highlights areas that warrant future study.
Figures

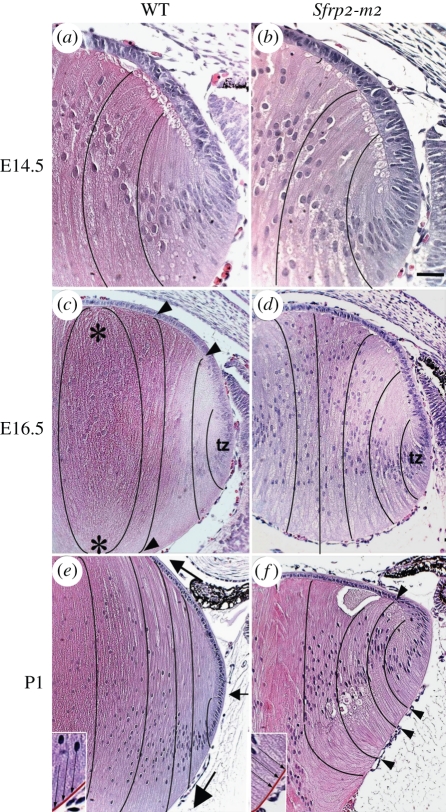
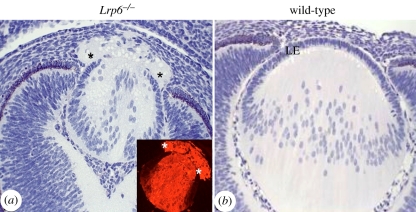
References
-
- McAvoy J. W. 1978. Cell division, cell elongation and distribution of alpha-, beta- and gamma-crystallins in the rat lens. J. Embryol. Exp. Morphol. 44, 149–165 - PubMed
-
- Coulombre J. L., Coulombre A. J. 1963. Lens development: fiber elongation and lens orientation. Science 142, 1489–1490 10.1126/science.142.3598.1489 (doi:10.1126/science.142.3598.1489) - DOI - PubMed
-
- Yamamoto Y. 1976. Growth of lens and ocular environment: role of neural retina in the growth of mouse lens as revealed by an implantation experiment. Dev. Growth Differ. 18, 273–278 10.1111/j.1440-169X.1976.00273.x (doi:10.1111/j.1440-169X.1976.00273.x) - DOI - PubMed
-
- Lovicu F. J., McAvoy J. W. 2005. Growth factor regulation of lens development. Dev. Biol. 280, 1–14 10.1016/j.ydbio.2005.01.020 (doi:10.1016/j.ydbio.2005.01.020) - DOI - PubMed
-
- West-Mays J. A., Pino G., Lovicu F. J. 2009. Development and use of the lens epithelial explant system to study lens differentiation and cataractogenesis. Prog. Retin. Eye Res. 29, 135–143 10.1016/j.preteyeres.2009.12.001 (doi:10.1016/j.preteyeres.2009.12.001) - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources