

Biological glass: structural determinants of eye lens transparency
- PMID: 21402584
- PMCID: PMC3061108
- DOI: 10.1098/rstb.2010.0302
Biological glass: structural determinants of eye lens transparency
Abstract
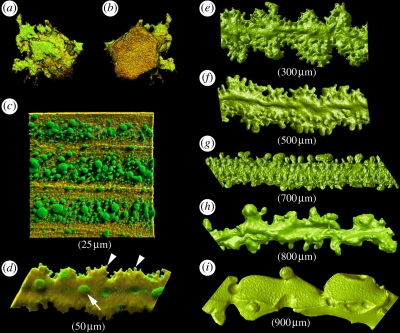
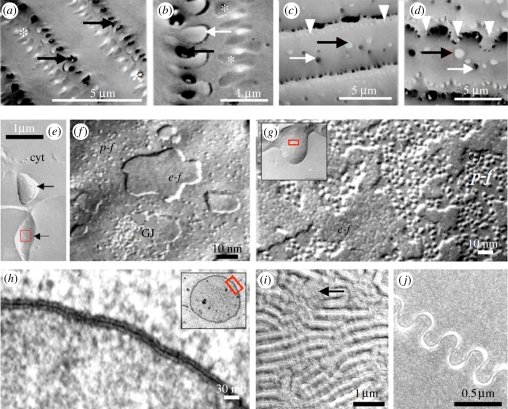
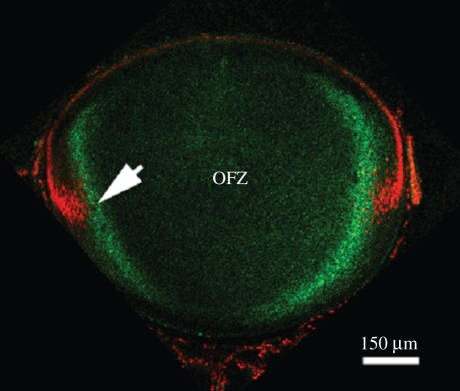
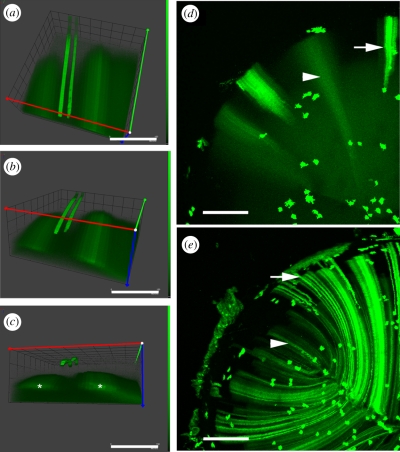
The purpose of the lens is to project a sharply focused, undistorted image of the visual surround onto the neural retina. The first pre-requisite, therefore, is that the tissue should be transparent. Despite the presence of remarkably high levels of protein, the lens cytosol remains transparent as a result of short-range-order interactions between the proteins. At a cellular level, the programmed elimination of nuclei and other light-scattering organelles from cells located within the pupillary space contributes directly to tissue transparency. Scattering at the cell borders is minimized by the close apposition of lens fibre cells facilitated by a plethora of adhesive proteins, some expressed only in the lens. Similarly, refractive index matching between lens membranes and cytosol is believed to minimize scatter. Refractive index matching between the cytoplasm of adjacent cells is achieved through the formation of cellular fusions that allow the intermingling of proteins. Together, these structural adaptations serve to minimize light scatter and enable this living, cellular structure to function as 'biological glass'.
Figures





References
-
- Michael R., Bron A. J. 2011. The ageing lens and cataract: a model of normal and pathological ageing. Phil. Trans. R. Soc. B 366, 1278–1292 10.1098/rstb.2010.0300 (doi:10.1098/rstb.2010.0300) - DOI - PMC - PubMed
-
- Muller L. J., Pels E., Schurmans L. R. H. M., Vrensen G. F. J. M. 2004. A new three-dimensional model of the organization of proteoglycans and collagen fibrils in the human corneal stroma. Exp. Eye Res. 78, 493–501 10.1016/S0014-4835(03)00206-9 (doi:10.1016/S0014-4835(03)00206-9) - DOI - PubMed
-
- Muller L. J., Pels L., Vrensen G. F. 1995. Novel aspects of the ultrastructural organization of human corneal keratocytes. Invest. Ophthalmol. Vis. Sci. 36, 2557–2567 - PubMed
-
- Kasthurirangan S., Markwell E. L., Atchison D. A., Pope J. M. 2008. In vivo study of changes in refractive index distribution in the human crystalline lens with age and accommodation. Invest. Ophthalmol. Vis. Sci. 49, 2531–2540 10.1167/iovs.07-1443 (doi:10.1167/iovs.07-1443) - DOI - PubMed
-
- Duncan M. K., Cvekl A., Kantorow M., Piatigorsky J. 2004. Lens crystallins. In Development of the ocular lens (eds Lovicu F. J., Robinson M. L.), pp. 119–150 Cambridge, UK: Cambridge University Press
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources