

Control of bone formation by the serpentine receptor Frizzled-9
- PMID: 21402791
- PMCID: PMC3063134
- DOI: 10.1083/jcb.201008012
Control of bone formation by the serpentine receptor Frizzled-9
Abstract
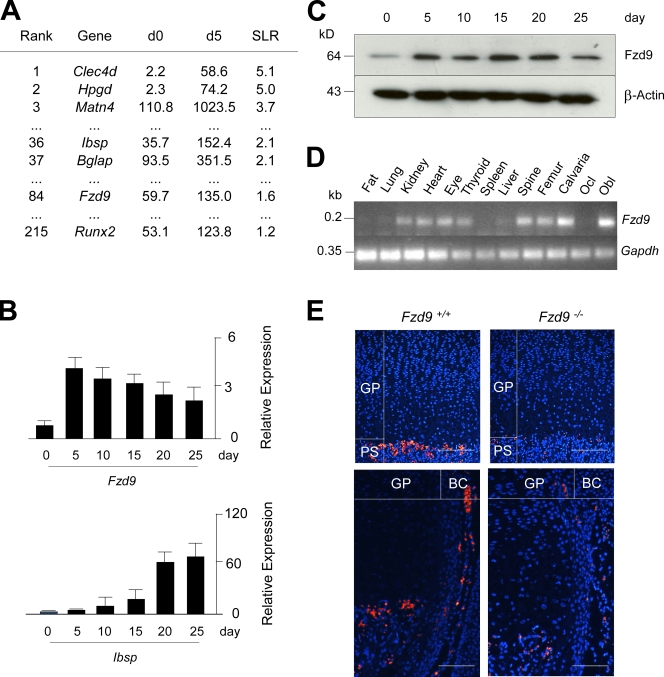
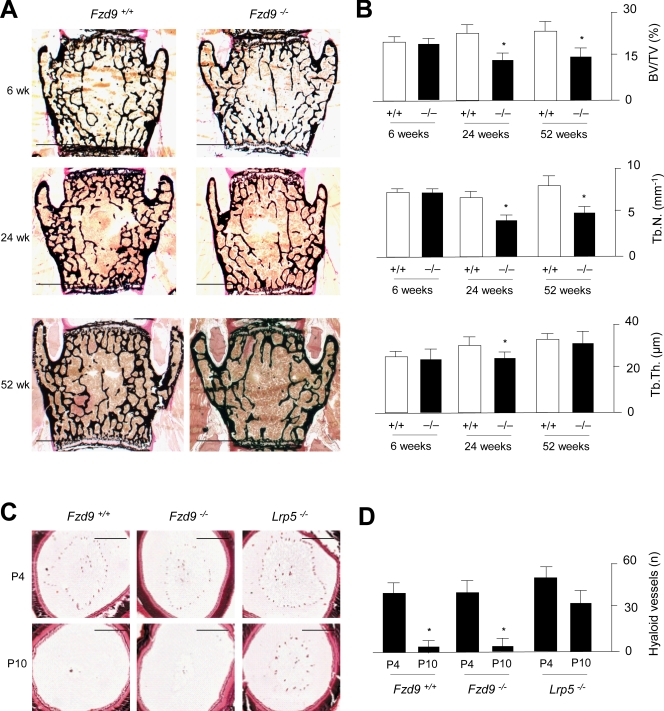
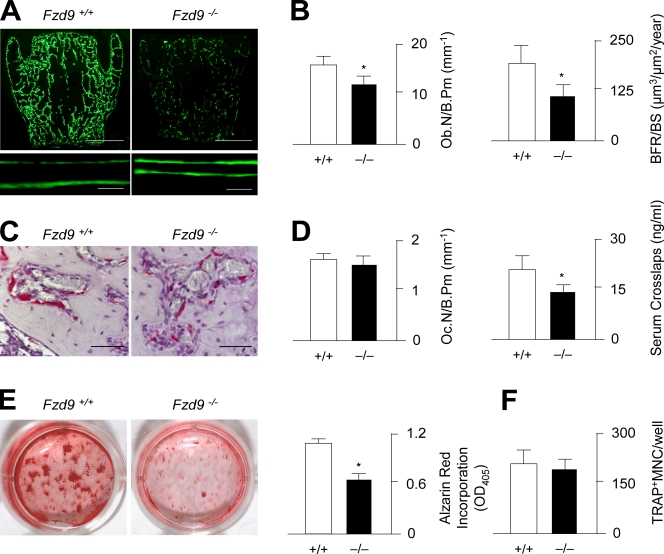
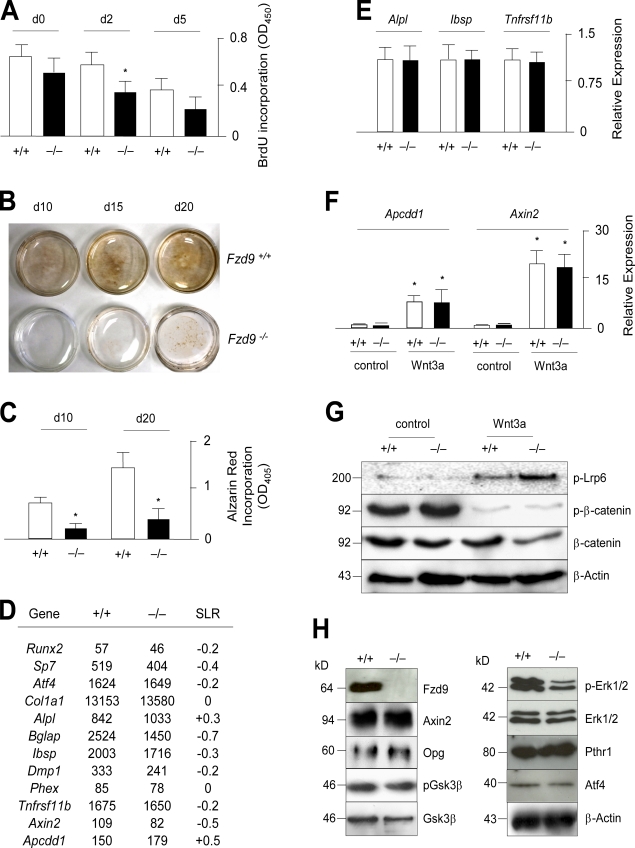
Although Wnt signaling in osteoblasts is of critical importance for the regulation of bone remodeling, it is not yet known which specific Wnt receptors of the Frizzled family are functionally relevant in this process. In this paper, we show that Fzd9 is induced upon osteoblast differentiation and that Fzd9(-/-) mice display low bone mass caused by impaired bone formation. Our analysis of Fzd9(-/-) primary osteoblasts demonstrated defects in matrix mineralization in spite of normal expression of established differentiation markers. In contrast, we observed a reduced expression of chemokines and interferon-regulated genes in Fzd9(-/-) osteoblasts. We also identified the ubiquitin-like modifier Isg15 as one potential downstream mediator of Fzd9 in these cells. Importantly, our molecular analysis further revealed that canonical Wnt signaling is not impaired in the absence of Fzd9, thus explaining the absence of a bone resorption phenotype. Collectively, our results reveal a previously unknown function of Fzd9 in osteoblasts, a finding that may have therapeutic implications for bone loss disorders.
Figures
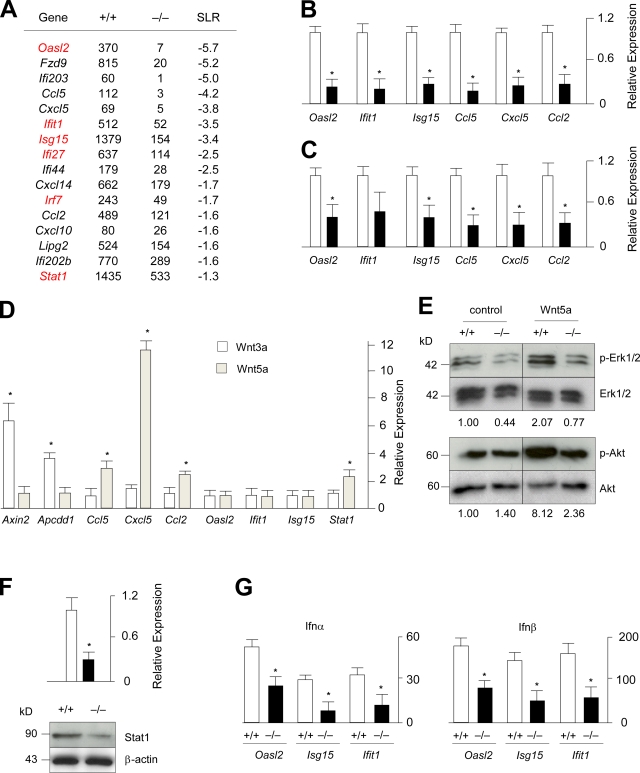
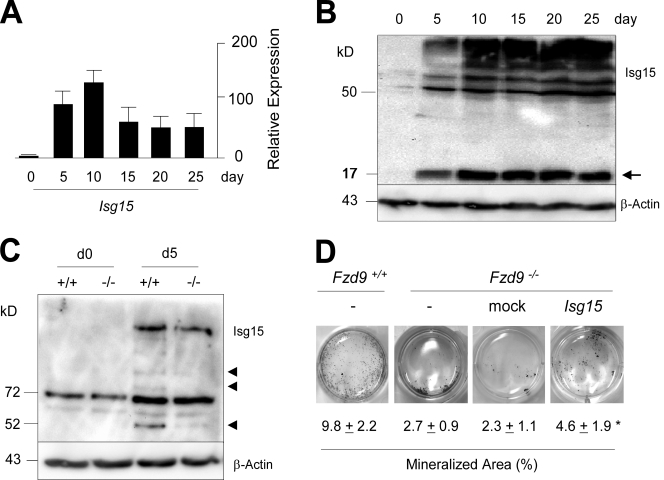
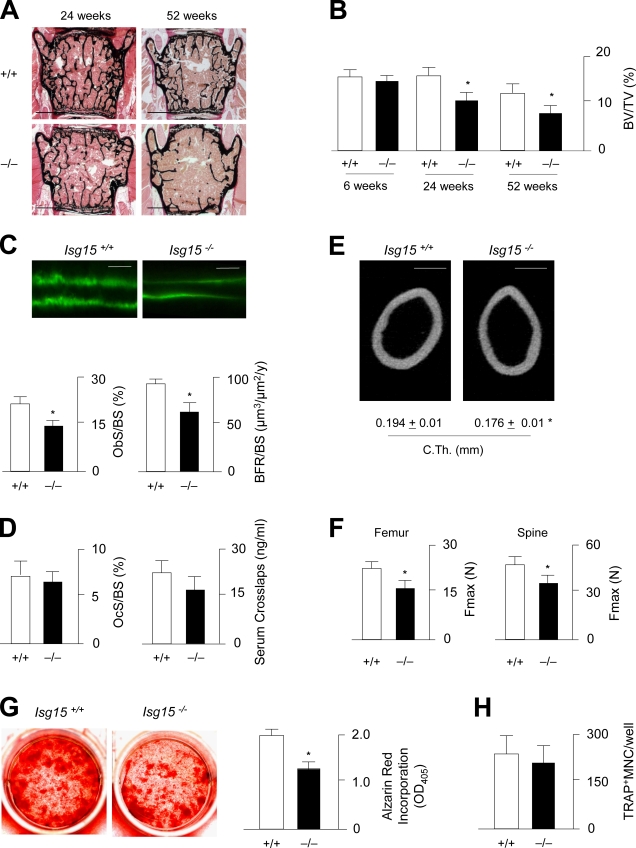
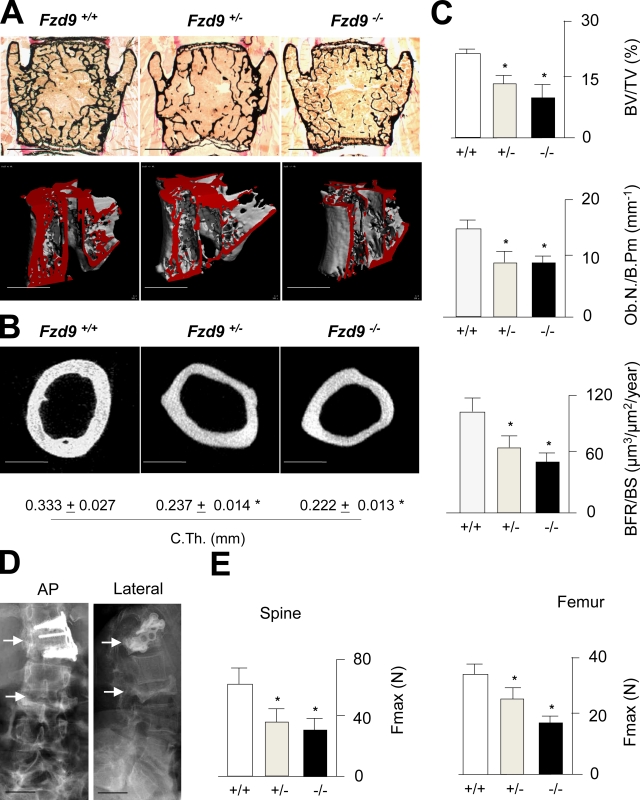
Comment in
-
Developement: Strong bones: got FZD9?Nat Rev Mol Cell Biol. 2011 May;12(5):280. doi: 10.1038/nrm3103. Epub 2011 Apr 7. Nat Rev Mol Cell Biol. 2011. PMID: 21471999 No abstract available.
References
-
- Almeida M., Han L., Bellido T., Manolagas S.C., Kousteni S. 2005. Wnt proteins prevent apoptosis of both uncommitted osteoblast progenitors and differentiated osteoblasts by beta-catenin-dependent and -independent signaling cascades involving Src/ERK and phosphatidylinositol 3-kinase/AKT. J. Biol. Chem. 280:41342–41351 10.1074/jbc.M502168200 - DOI - PubMed
-
- Bodine P.V., Zhao W., Kharode Y.P., Bex F.J., Lambert A.J., Goad M.B., Gaur T., Stein G.S., Lian J.B., Komm B.S. 2004. The Wnt antagonist secreted frizzled-related protein-1 is a negative regulator of trabecular bone formation in adult mice. Mol. Endocrinol. 18:1222–1237 10.1210/me.2003-0498 - DOI - PubMed
-
- Cummings S.R., San Martin J., McClung M.R., Siris E.S., Eastell R., Reid I.R., Delmas P., Zoog H.B., Austin M., Wang A., et al. ; FREEDOM Trial 2009. Denosumab for prevention of fractures in postmenopausal women with osteoporosis. N. Engl. J. Med. 361:756–765 (published erratum appears in N. Engl. J. Med. 2009. 361: 1914) 10.1056/NEJMoa0809493 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
