

Transcriptional coupling of neighboring genes and gene expression noise: evidence that gene orientation and noncoding transcripts are modulators of noise
- PMID: 21402863
- PMCID: PMC5654408
- DOI: 10.1093/gbe/evr025
Transcriptional coupling of neighboring genes and gene expression noise: evidence that gene orientation and noncoding transcripts are modulators of noise
Abstract
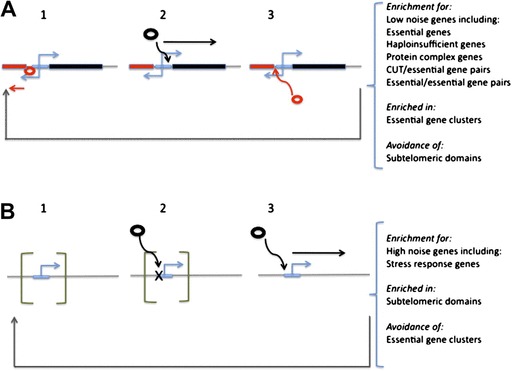
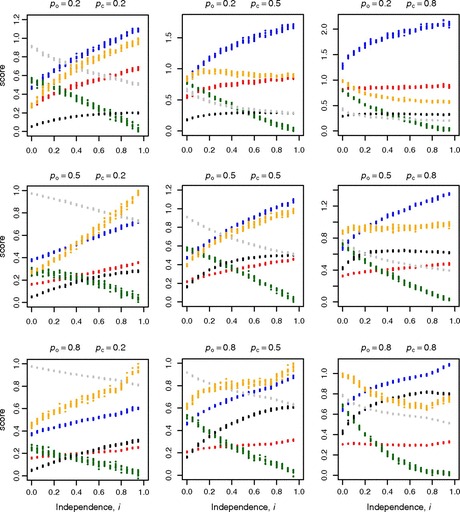
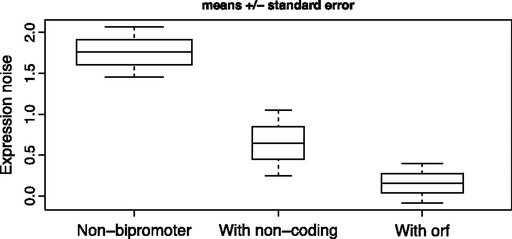
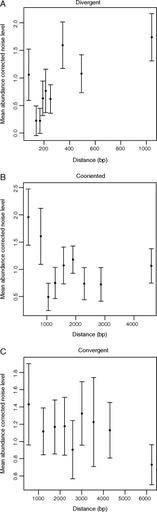
How is noise in gene expression modulated? Do mechanisms of noise control impact genome organization? In yeast, the expression of one gene can affect that of a very close neighbor. As the effect is highly regionalized, we hypothesize that genes in different orientations will have differing degrees of coupled expression and, in turn, different noise levels. Divergently organized gene pairs, in particular those with bidirectional promoters, have close promoters, maximizing the likelihood that expression of one gene affects the neighbor. With more distant promoters, the same is less likely to hold for gene pairs in nondivergent orientation. Stochastic models suggest that coupled chromatin dynamics will typically result in low abundance-corrected noise (ACN). Transcription of noncoding RNA (ncRNA) from a bidirectional promoter, we thus hypothesize to be a noise-reduction, expression-priming, mechanism. The hypothesis correctly predicts that protein-coding genes with a bidirectional promoter, including those with a ncRNA partner, have lower ACN than other genes and divergent gene pairs uniquely have correlated ACN. Moreover, as predicted, ACN increases with the distance between promoters. The model also correctly predicts ncRNA transcripts to be often divergently transcribed from genes that a priori would be under selection for low noise (essential genes, protein complex genes) and that the latter genes should commonly reside in divergent orientation. Likewise, that genes with bidirectional promoters are rare subtelomerically, cluster together, and are enriched in essential gene clusters is expected and observed. We conclude that gene orientation and transcription of ncRNAs are candidate modulators of noise.
Figures
References
-
- Balaji S, Babu MM, Iyer LM, Luscombe NM, Aravind L. Comprehensive analysis of combinatorial regulation using the transcriptional regulatory network of yeast. J Mol Biol. 2006;360:213–227. - PubMed
-
- Bar-Even A, et al. Noise in protein expression scales with natural protein abundance. Nat Genet. 2006;38:636–643. - PubMed
-
- Basehoar AD, Zanton SJ, Pugh BF. Identification and distinct regulation of yeast TATA box-containing genes. Cell. 2004;116:699–709. - PubMed
-
- Batada NN, Hurst LD. Evolution of chromosome organization driven by selection for reduced gene expression noise. Nat Genet. 2007;39:945–949. - PubMed
-
- Batada NN, Urrutia AO, Hurst LD. Chromatin remodelling is a major source of coexpression of linked genes in yeast. Trends Genet. 2007;23:480–484. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
