

Identity, regulation, and activity of inducible diterpenoid phytoalexins in maize
- PMID: 21402917
- PMCID: PMC3069164
- DOI: 10.1073/pnas.1014714108
Identity, regulation, and activity of inducible diterpenoid phytoalexins in maize
Abstract
Phytoalexins constitute a broad category of pathogen- and insect-inducible biochemicals that locally protect plant tissues. Because of their agronomic significance, maize and rice have been extensively investigated for their terpenoid-based defenses, which include insect-inducible monoterpene and sesquiterpene volatiles. Rice also produces a complex array of pathogen-inducible diterpenoid phytoalexins. Despite the demonstration of fungal-induced ent-kaur-15-ene production in maize over 30 y ago, the identity of functionally analogous maize diterpenoid phytoalexins has remained elusive. In response to stem attack by the European corn borer (Ostrinia nubilalis) and fungi, we observed the induced accumulation of six ent-kaurane-related diterpenoids, collectively termed kauralexins. Isolation and identification of the predominant Rhizopus microsporus-induced metabolites revealed ent-kaur-19-al-17-oic acid and the unique analog ent-kaur-15-en-19-al-17-oic acid, assigned as kauralexins A3 and B3, respectively. Encoding an ent-copalyl diphosphate synthase, fungal-induced An2 transcript accumulation precedes highly localized kauralexin production, which can eventually exceed 100 μg · g(-1) fresh weight. Pharmacological applications of jasmonic acid and ethylene also synergize the induced accumulation of kauralexins. Occurring at elevated levels in the scutella of all inbred lines examined, kauralexins appear ubiquitous in maize. At concentrations as low as 10 μg · mL(-1), kauralexin B3 significantly inhibited the growth of the opportunistic necrotroph R. microsporus and the causal agent of anthracnose stalk rot, Colletotrichum graminicola. Kauralexins also exhibited significant O. nubilalis antifeedant activity. Our work establishes the presence of diterpenoid defenses in maize and enables a more detailed analysis of their biosynthetic pathways, regulation, and crop defense function.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
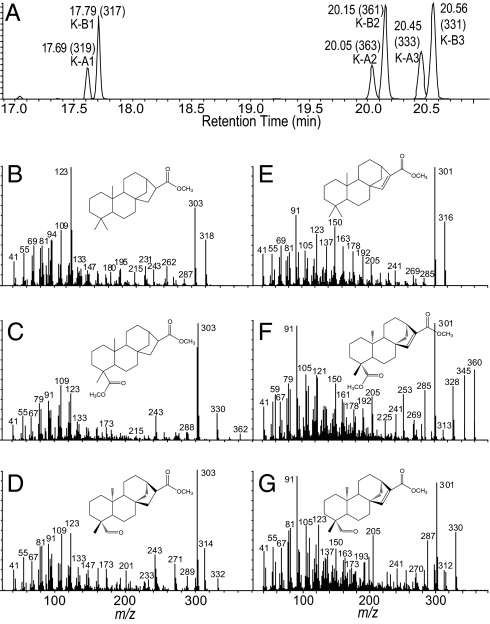
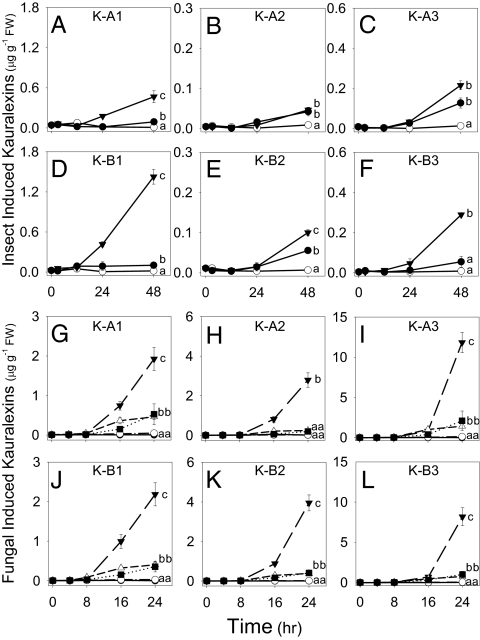
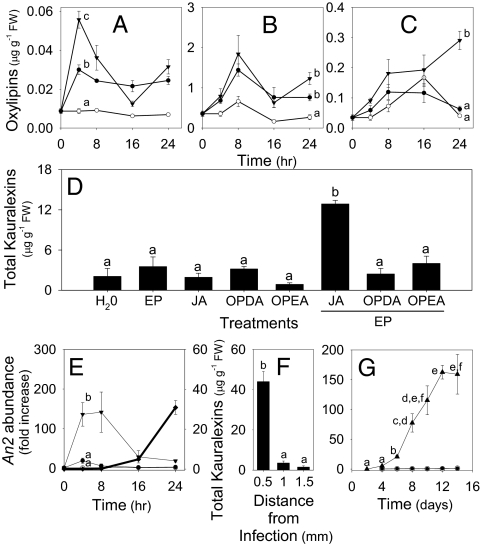
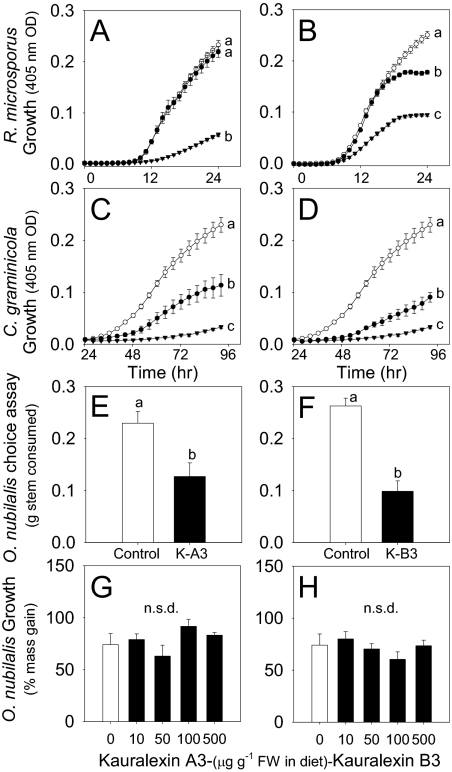
References
-
- Smith CJ. Accumulation of phytoalexins: Defence mechanism and stimulus response system. New Phytol. 1996;132:1–45. - PubMed
-
- Delannoy E, et al. Resistance of cotton towards Xanthomonas campestris pv. malvacearum. Annu Rev Phytopathol. 2005;43:63–82. - PubMed
-
- McAuslane HJ, Alborn HT. Systemic induction of allelochemicals in glanded and glandless isogenic cotton by Spodoptera exigua feeding. J Chem Ecol. 1998;24:399–416.
-
- Toyomasu T. Recent advances regarding diterpene cyclase genes in higher plants and fungi. Biosci Biotechnol Biochem. 2008;72:1168–1175. - PubMed
-
- Peters RJ. Uncovering the complex metabolic network underlying diterpenoid phytoalexin biosynthesis in rice and other cereal crop plants. Phytochemistry. 2006;67:2307–2317. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
