

Iron enzyme ribulose-5-phosphate 3-epimerase in Escherichia coli is rapidly damaged by hydrogen peroxide but can be protected by manganese
- PMID: 21402925
- PMCID: PMC3069151
- DOI: 10.1073/pnas.1100410108
Iron enzyme ribulose-5-phosphate 3-epimerase in Escherichia coli is rapidly damaged by hydrogen peroxide but can be protected by manganese
Abstract
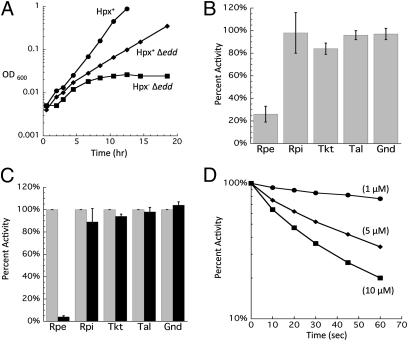
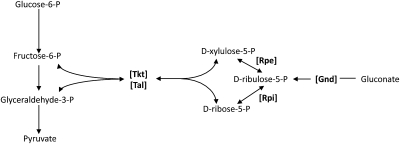
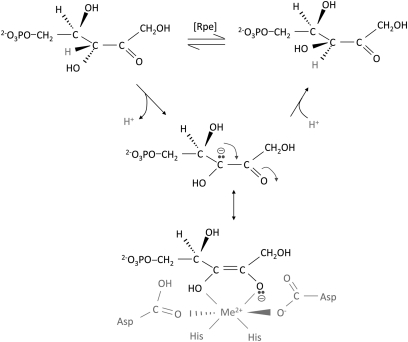
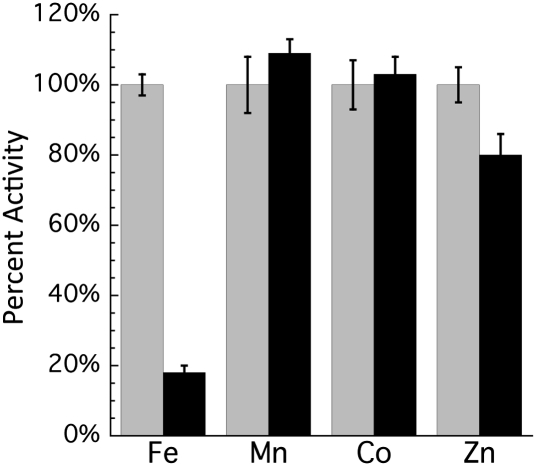
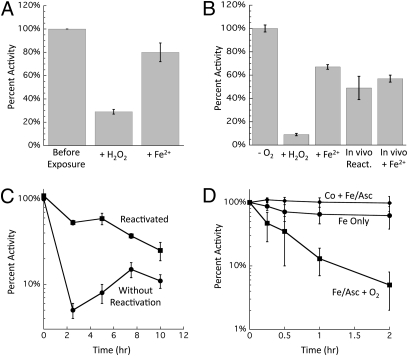
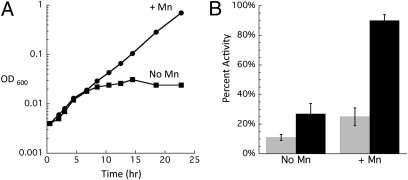
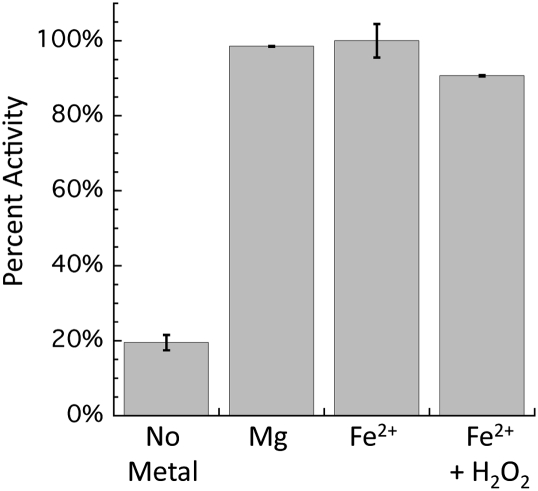
H(2)O(2) is commonly generated in biological habitats by environmental chemistry and by cellular immune responses. H(2)O(2) penetrates cells, disrupts metabolism, and blocks growth; it therefore is of interest to identify the major cellular molecules that H(2)O(2) damages and the strategies by which cells protect themselves from it. We used a strain of Escherichia coli that lacks catalases and peroxidases to impose protracted low-grade H(2)O(2) stress. Physiological analysis indicated that the pentose-phosphate pathway, in particular, was poisoned by submicromolar intracellular H(2)O(2). Assays determined that ribulose-5-phosphate 3-epimerase (Rpe) was specifically inactivated. In vitro studies demonstrated that Rpe employs a ferrous iron atom as a solvent-exposed cofactor and that H(2)O(2) rapidly oxidizes this metal in a Fenton reaction. The oxidized iron is released immediately, causing a loss of activity. Most Rpe proteins could be reactivated by remetallation; however, a small fraction of proteins were irreversibly damaged by each oxidation cycle, and so repeated cycles of oxidation and remetallation progressively led to permanent inactivation of the entire Rpe pool. Manganese import and iron sequestration are key elements of the H(2)O(2) stress response, and we found that manganese can activate Rpe in vitro in place of iron, converting the enzyme to a form that is unaffected by H(2)O(2). Indeed, the provision of manganese to H(2)O(2)-stressed cells protected Rpe and enabled the pentose-phosphate pathway to retain function. These data indicate that mononuclear iron enzymes can be primary targets of H(2)O(2) stress and that cells adapt by shifting from iron- to manganese-centered metabolism.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Loew O. A new enzyme of general occurrence in organisms. Science. 1900;11:701–702. - PubMed
-
- Glass GA, et al. The respiratory burst oxidase of human neutrophils. Further studies of the purified enzyme. J Biol Chem. 1986;261:13247–13251. - PubMed
-
- Carmel-Harel O, Storz G. Roles of the glutathione- and thioredoxin-dependent reduction systems in the Escherichia coli and Saccharomyces cerevisiae responses to oxidative stress. Annu Rev Microbiol. 2000;54:439–461. - PubMed
-
- Lee JW, Helmann JD. The PerR transcription factor senses H2O2 by metal-catalysed histidine oxidation. Nature. 2006;440:363–367. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
