

Intestinal epithelial cells as producers but not targets of chronic TNF suffice to cause murine Crohn-like pathology
- PMID: 21402942
- PMCID: PMC3069201
- DOI: 10.1073/pnas.1007811108
Intestinal epithelial cells as producers but not targets of chronic TNF suffice to cause murine Crohn-like pathology
Abstract
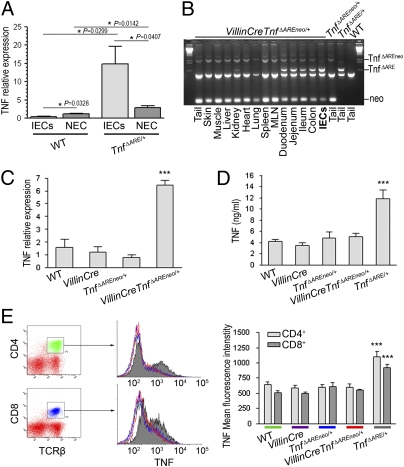
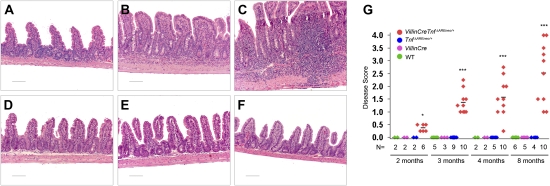
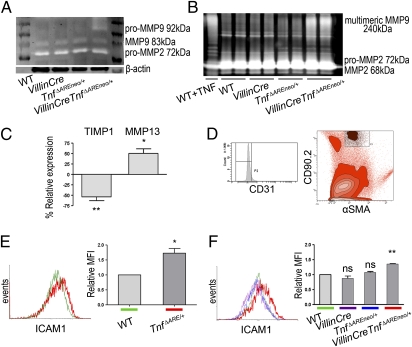
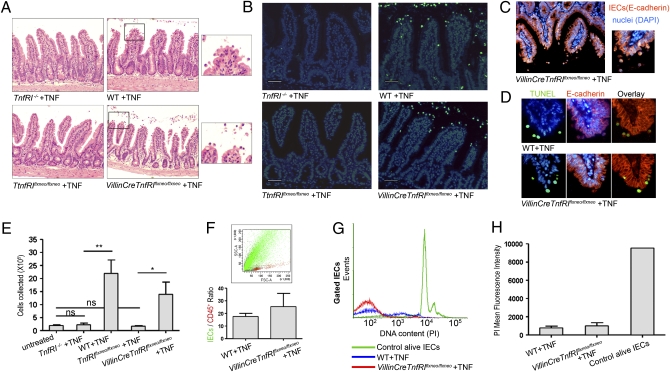
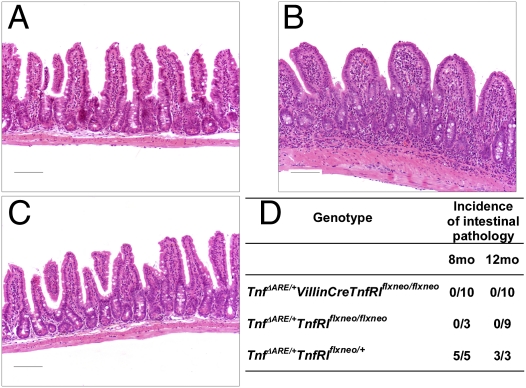
TNF plays a crucial role in the pathogenesis of Crohn disease. Dysregulated TNF production in mice that bear the genetic deletion of the TNF AU-rich regulatory elements (ARE) (Tnf(ΔARE/+) mice) results in TNF receptor I (TNFRI)-dependent spontaneous Crohn-like pathology. Current concepts consider intestinal epithelial cell (IEC) responses to TNF to be critical for intestinal pathology, but the potential contribution of IEC-derived TNF in disease pathogenesis has not been addressed. In this study we examined whether IEC are sufficient as cellular targets or sources of TNF in the development of intestinal pathology. Using IEC-specific reactivation of a hypomorphic Tnf(ΔAREneo) allele in mice, we show that selective chronic overproduction of TNF by IEC suffices to cause full development of Crohn-like pathology. Epithelial TNF overexpression leads to early activation of the underlying intestinal myofibroblast, a cell type previously identified as a sufficient target of TNF for disease development in the Tnf(ΔARE) model. By contrast, restricted TNFRI expression on IEC although sufficient to confer IEC apoptosis after acute exogenous TNF administration, fails to induce pathology following chronic specific targeting of IEC by endogenous TNF in Tnf(ΔARE/+) mice. Our results argue against IEC being early and sufficient responders to chronic TNF-mediated pathogenic signals and suggest that proinflammatory aberrations leading to chronic TNF production by IEC may initiate pathology in Crohn disease.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Intestinal-specific TNFα overexpression induces Crohn's-like ileitis in mice.PLoS One. 2013 Aug 20;8(8):e72594. doi: 10.1371/journal.pone.0072594. eCollection 2013. PLoS One. 2013. PMID: 23977323 Free PMC article.
-
FADD prevents RIP3-mediated epithelial cell necrosis and chronic intestinal inflammation.Nature. 2011 Jul 31;477(7364):330-4. doi: 10.1038/nature10273. Nature. 2011. PMID: 21804564
-
Herbs-partitioned moxibustion alleviates aberrant intestinal epithelial cell apoptosis by upregulating A20 expression in a mouse model of Crohn's disease.World J Gastroenterol. 2019 May 7;25(17):2071-2085. doi: 10.3748/wjg.v25.i17.2071. World J Gastroenterol. 2019. PMID: 31114134 Free PMC article.
-
p53 mediates TNF-induced epithelial cell apoptosis in IBD.Am J Pathol. 2012 Oct;181(4):1306-15. doi: 10.1016/j.ajpath.2012.06.016. Epub 2012 Aug 3. Am J Pathol. 2012. PMID: 22863952 Free PMC article.
-
Dysregulation of Intestinal Epithelial Cell RIPK Pathways Promotes Chronic Inflammation in the IBD Gut.Front Immunol. 2019 May 20;10:1094. doi: 10.3389/fimmu.2019.01094. eCollection 2019. Front Immunol. 2019. PMID: 31164887 Free PMC article. Review.
Cited by
-
The Balance of TNF Mediated Pathways Regulates Inflammatory Cell Death Signaling in Healthy and Diseased Tissues.Front Cell Dev Biol. 2020 May 21;8:365. doi: 10.3389/fcell.2020.00365. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32671059 Free PMC article. Review.
-
HMGB1 released from intestinal epithelia damaged by cholera toxin adjuvant contributes to activation of mucosal dendritic cells and induction of intestinal cytotoxic T lymphocytes and IgA.Cell Death Dis. 2018 May 24;9(6):631. doi: 10.1038/s41419-018-0665-z. Cell Death Dis. 2018. PMID: 29795370 Free PMC article.
-
Redeeming an old foe: protective as well as pathophysiological roles for tumor necrosis factor in inflammatory bowel disease.Am J Physiol Gastrointest Liver Physiol. 2015 Feb 1;308(3):G161-70. doi: 10.1152/ajpgi.00142.2014. Epub 2014 Dec 4. Am J Physiol Gastrointest Liver Physiol. 2015. PMID: 25477373 Free PMC article. Review.
-
The transition of acute to chronic bowel inflammation in spondyloarthritis.Nat Rev Rheumatol. 2012 Apr 17;8(5):288-95. doi: 10.1038/nrrheum.2012.42. Nat Rev Rheumatol. 2012. PMID: 22508429 Review.
-
When Insult Is Added to Injury: Cross Talk between ILCs and Intestinal Epithelium in IBD.Mediators Inflamm. 2016;2016:9765238. doi: 10.1155/2016/9765238. Epub 2016 Aug 4. Mediators Inflamm. 2016. PMID: 27578924 Free PMC article. Review.
References
-
- Xavier RJ, Podolsky DK. Unravelling the pathogenesis of inflammatory bowel disease. Nature. 2007;448:427–434. - PubMed
-
- Podolsky DK. Inflammatory bowel disease. N Engl J Med. 2002;347:417–429. - PubMed
-
- Cho JH. The genetics and immunopathogenesis of inflammatory bowel disease. Nat Rev Immunol. 2008;8:458–466. - PubMed
-
- Strober W, Fuss IJ, Blumberg RS. The immunology of mucosal models of inflammation. Annu Rev Immunol. 2002;20:495–549. - PubMed
-
- Targan SR, et al. Crohn's Disease cA2 Study Group. A short-term study of chimeric monoclonal antibody cA2 to tumor necrosis factor alpha for Crohn's disease. N Engl J Med. 1997;337:1029–1035. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
