

Histone methyltransferase EZH2 induces Akt-dependent genomic instability and BRCA1 inhibition in breast cancer
- PMID: 21406404
- PMCID: PMC3071296
- DOI: 10.1158/0008-5472.CAN-10-1933
Histone methyltransferase EZH2 induces Akt-dependent genomic instability and BRCA1 inhibition in breast cancer
Abstract
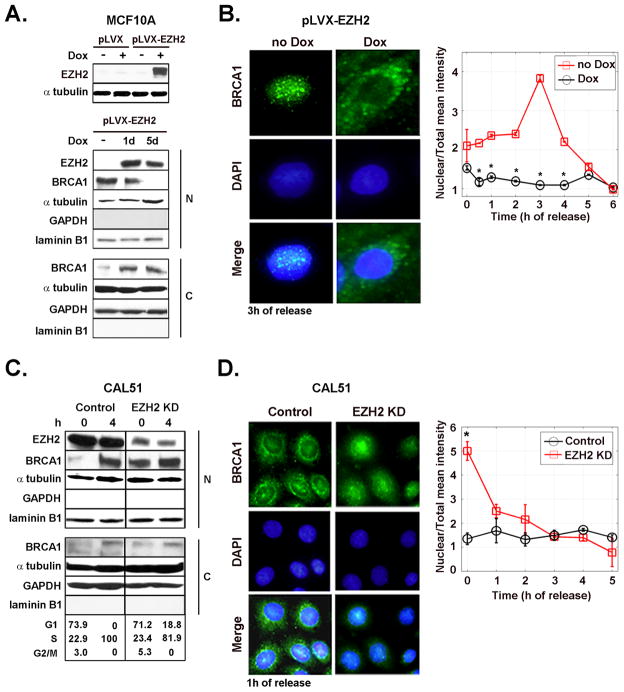
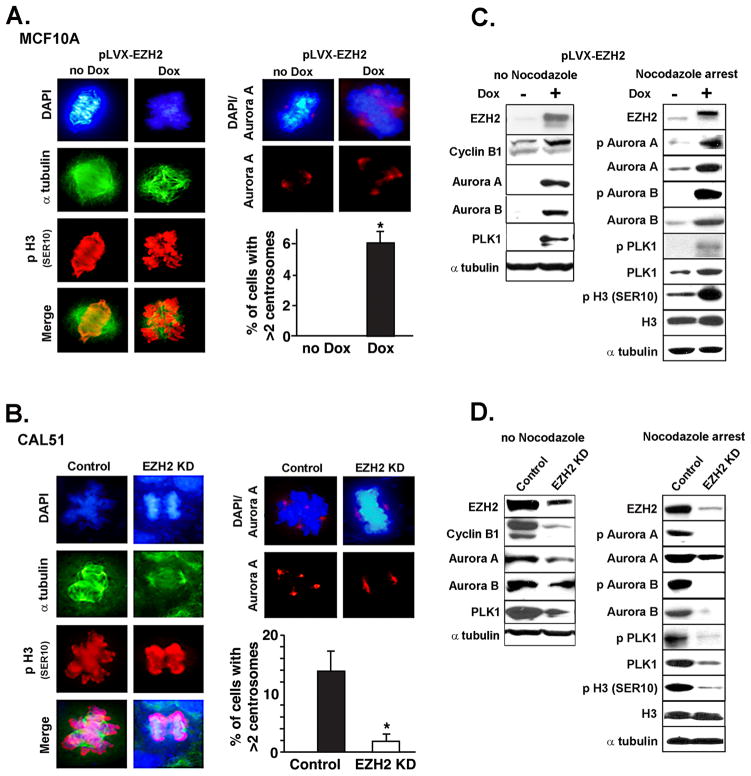
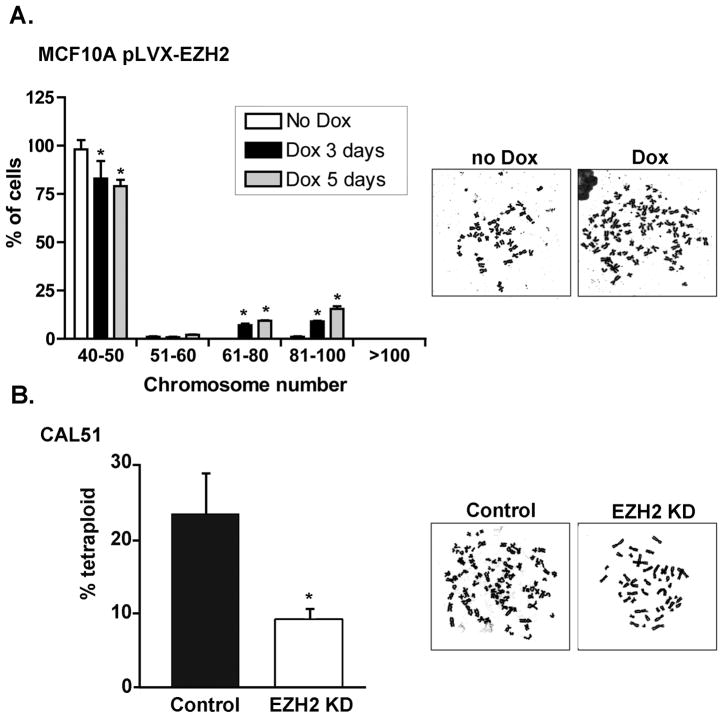
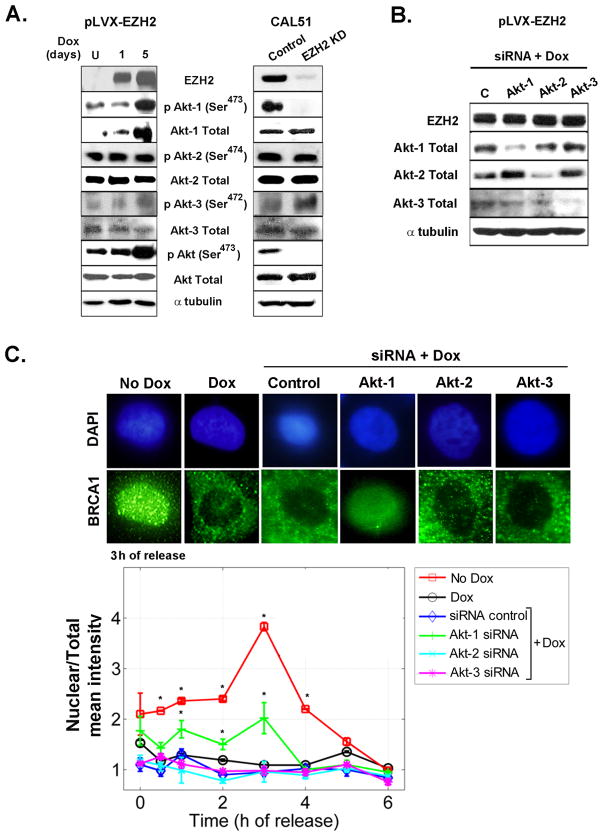
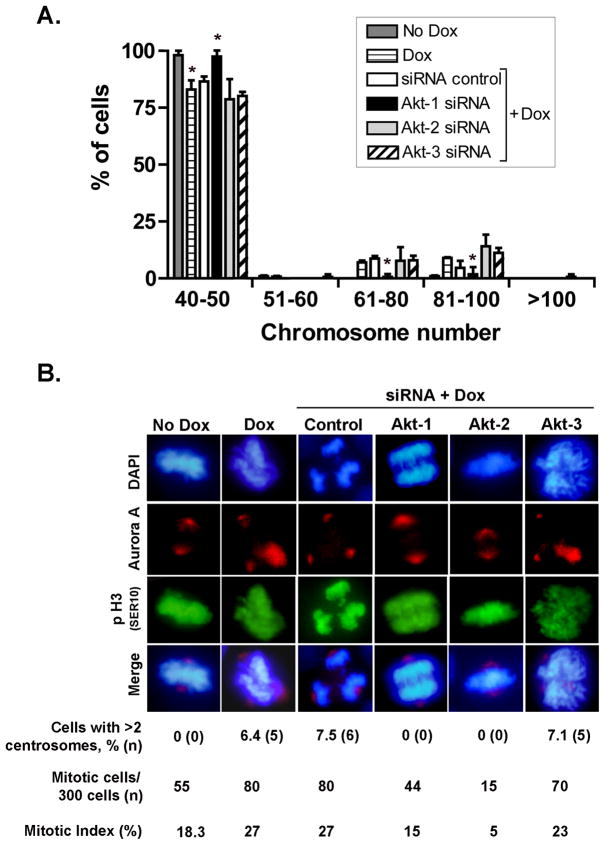
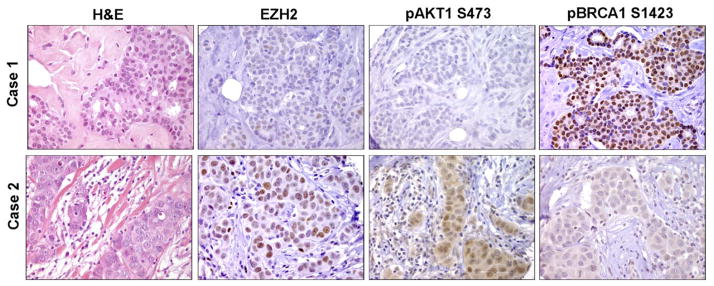
Increased levels of EZH2, a critical regulator of cellular memory, signal the presence of metastasis and poor outcome in breast cancer patients. High levels of EZH2 are associated with nuclear pleomorphism, lack of estrogen receptor expression, and decreased nuclear levels of BRCA1 tumor suppressor protein in invasive breast carcinomas. The mechanism by which EZH2 overexpression promotes the growth of poorly differentiated invasive carcinomas remains to be defined. Here, we show that EZH2 controls the intracellular localization of BRCA1 protein. Conditional doxycycline-induced upregulation of EZH2 in benign mammary epithelial cells results in nuclear export of BRCA1 protein, aberrant mitoses with extra centrosomes, and genomic instability. EZH2 inhibition in CAL51 breast cancer cells induces BRCA1 nuclear localization and rescues defects in ploidy and mitosis. Mechanistically, EZH2 overexpression is sufficient for activation of the phosphoinositide 3-kinase/Akt (PI3K/Akt) pathway specifically through activation of Akt isoform 1. EZH2-induced BRCA1 nuclear export, aneuploidy, and mitotic defects were prevented by treatment with the PI3K inhibitors LY294002 or wortmannin. Targeted inhibition of Akt-1, Akt-2, and Akt-3 isoforms revealed that the EZH2-induced phenotype requires specific activation of Akt-1. The relevance of our studies to human breast cancer is highlighted by the finding that high EZH2 protein levels are associated with upregulated expression of phospho-Akt-1 (Ser473) and decreased nuclear expression of phospho-BRCA1 (Ser1423) in 39% of invasive breast carcinomas. These results enable us to pinpoint one mechanism by which EZH2 regulates BRCA1 expression and genomic stability mediated by the PI3K/Akt-1 pathway.
© 2011 AACR.
Figures
References
-
- Satijn DP, Otte AP. Polycomb group protein complexes: do different complexes regulate distinct target genes? Biochim Biophys Acta. 1999;1447:1–16. - PubMed
-
- Ringrose L, Paro R. Epigenetic regulation of cellular memory by the Polycomb and Trithorax group proteins. Annu Rev Genet. 2004;38:413–43. - PubMed
-
- Haupt Y, Alexander WS, Barri G, Klinken SP, Adams JM. Novel zinc finger gene implicated as myc collaborator by retrovirally accelerated lymphomagenesis in E mu-myc transgenic mice. Cell. 1991;65:753–63. - PubMed
-
- Hess JL. MLL: a histone methyltransferase disrupted in leukemia. Trends Mol Med. 2004;10:500–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
