

The multifunctional Ccr4-Not complex directly promotes transcription elongation
- PMID: 21406554
- PMCID: PMC3059832
- DOI: 10.1101/gad.2020911
The multifunctional Ccr4-Not complex directly promotes transcription elongation
Abstract
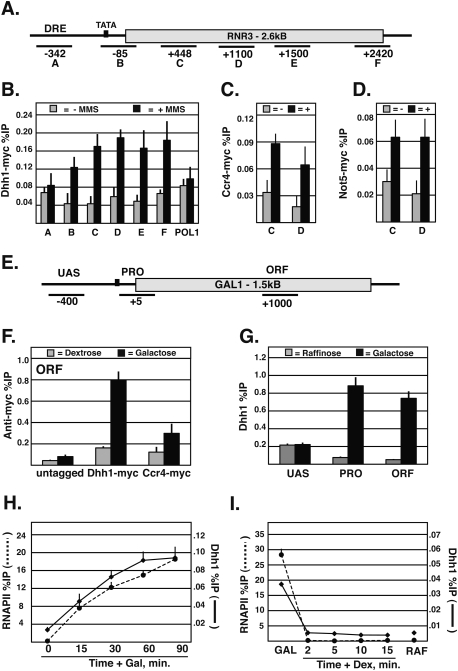
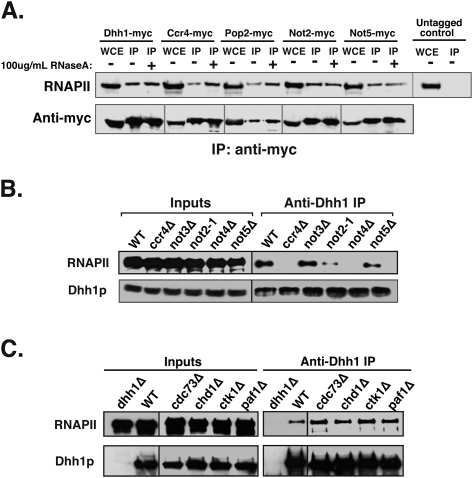
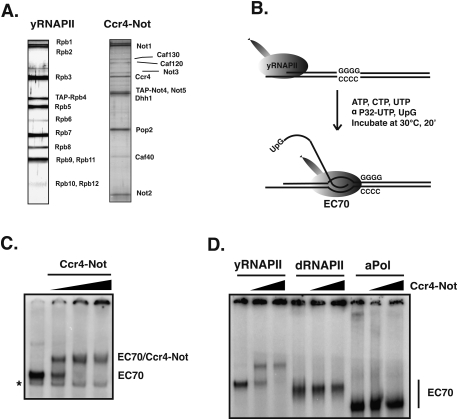
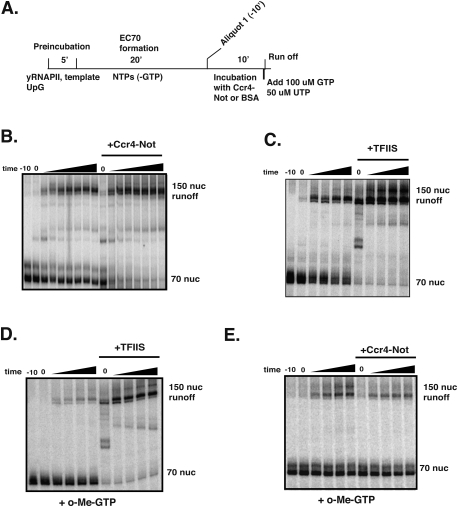
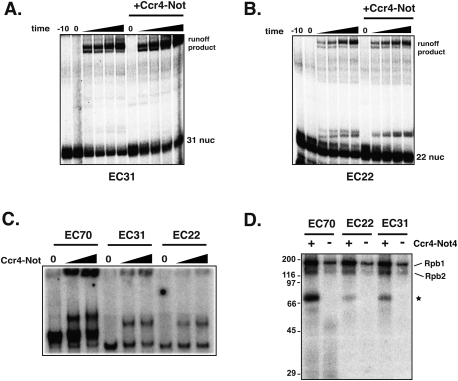
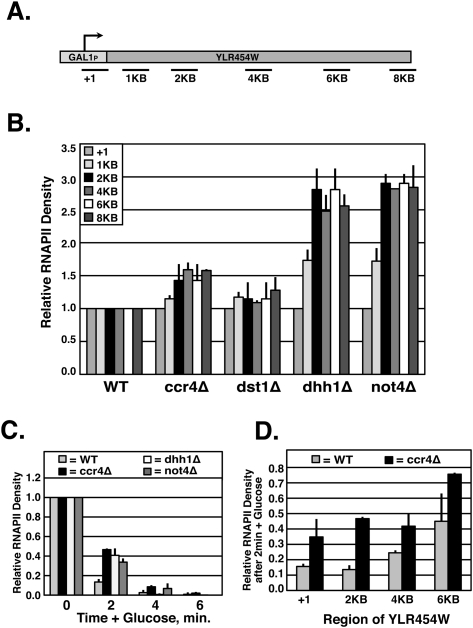
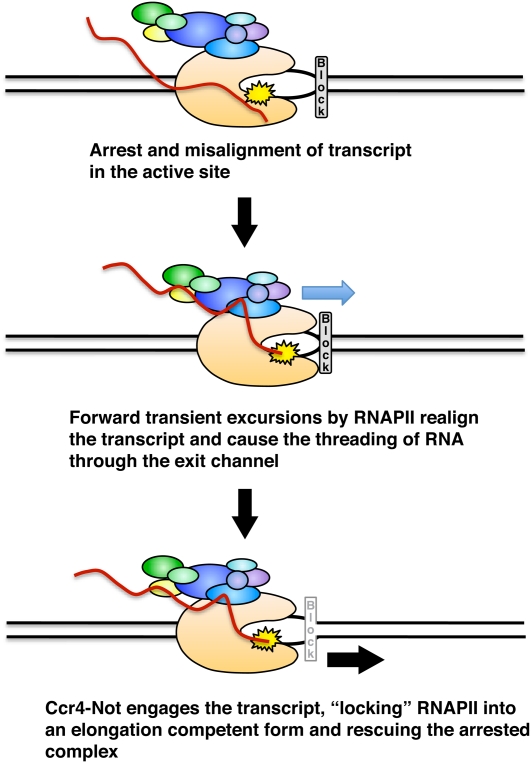
The Ccr4-Not complex has been implicated in the control of multiple steps of mRNA metabolism; however, its functions in transcription remain ambiguous. The discovery that Ccr4/Pop2 is the major cytoplasmic mRNA deadenylase and the detection of Not proteins within mRNA processing bodies have raised questions about the roles of the Ccr4-Not complex in transcription. Here we firmly establish Ccr4-Not as a positive elongation factor for RNA polymerase II (RNAPII). The Ccr4-Not complex is targeted to the coding region of genes in a transcription-dependent manner similar to RNAPII and promotes elongation in vivo. Furthermore, Ccr4-Not interacts directly with elongating RNAPII complexes and stimulates transcription elongation of arrested polymerase in vitro. Ccr4-Not can reactivate backtracked RNAPII using a mechanism different from that of the well-characterized elongation factor TFIIS. While not essential for its interaction with elongation complexes, Ccr4-Not interacts with the emerging transcript and promotes elongation in a manner dependent on transcript length, although this interaction is not required for it to bind RNAPII. Our comprehensive analysis shows that Ccr4-Not directly regulates transcription, and suggests it does so by promoting the resumption of elongation of arrested RNAPII when it encounters transcriptional blocks in vivo.
Figures
References
-
- Arndt KM, Kane CM 2003. Running with RNA polymerase: eukaryotic transcript elongation. Trends Genet 19: 543–550 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases