

Multiple facets of Arabidopsis seedling development require indole-3-butyric acid-derived auxin
- PMID: 21406624
- PMCID: PMC3082277
- DOI: 10.1105/tpc.111.083071
Multiple facets of Arabidopsis seedling development require indole-3-butyric acid-derived auxin
Abstract
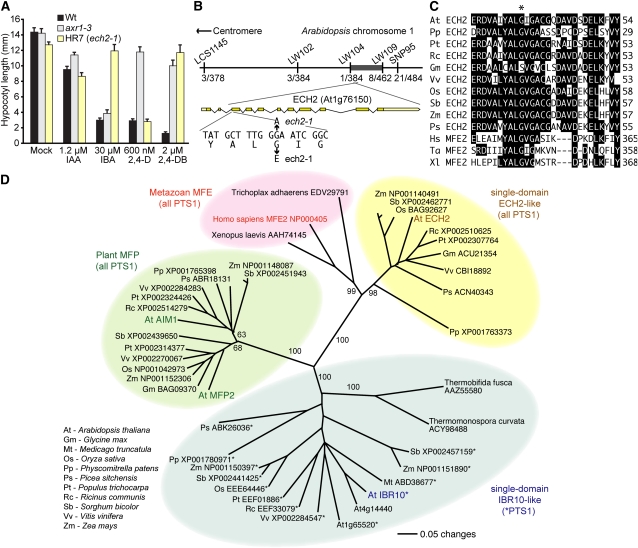
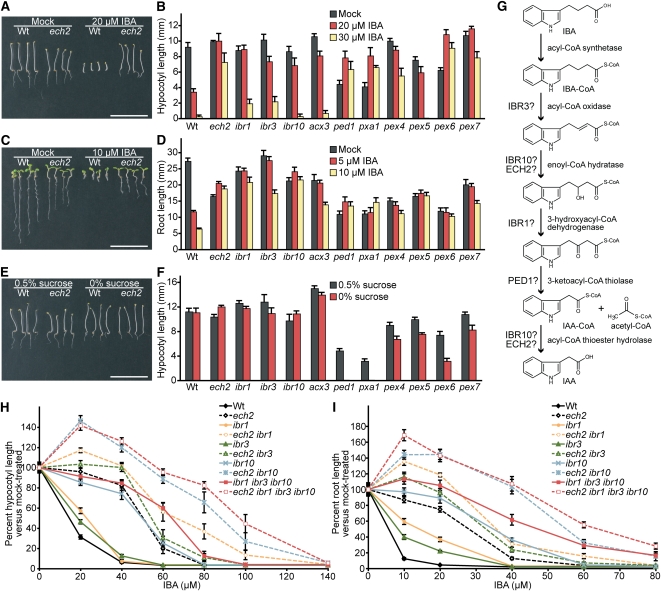
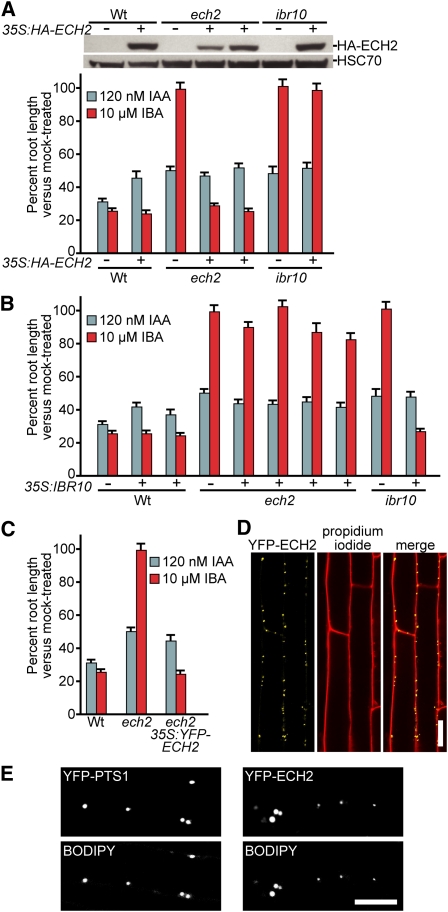
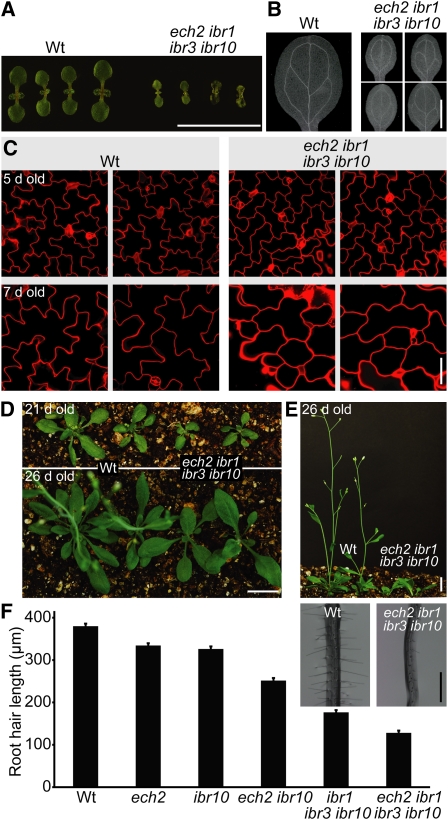
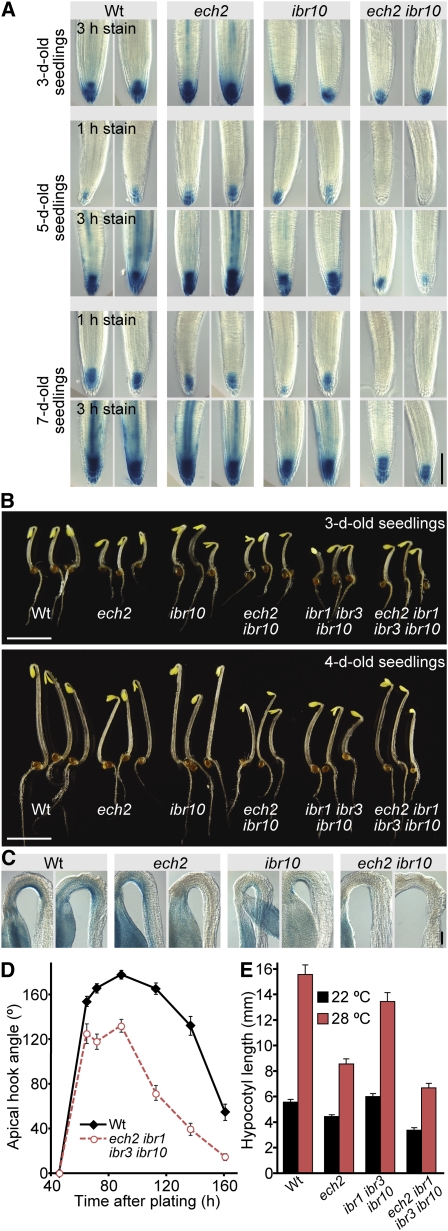
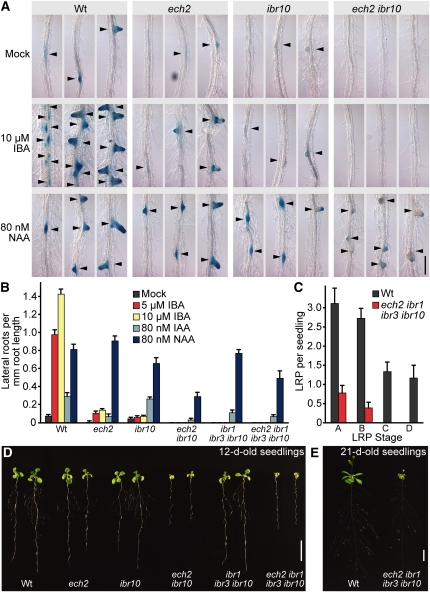
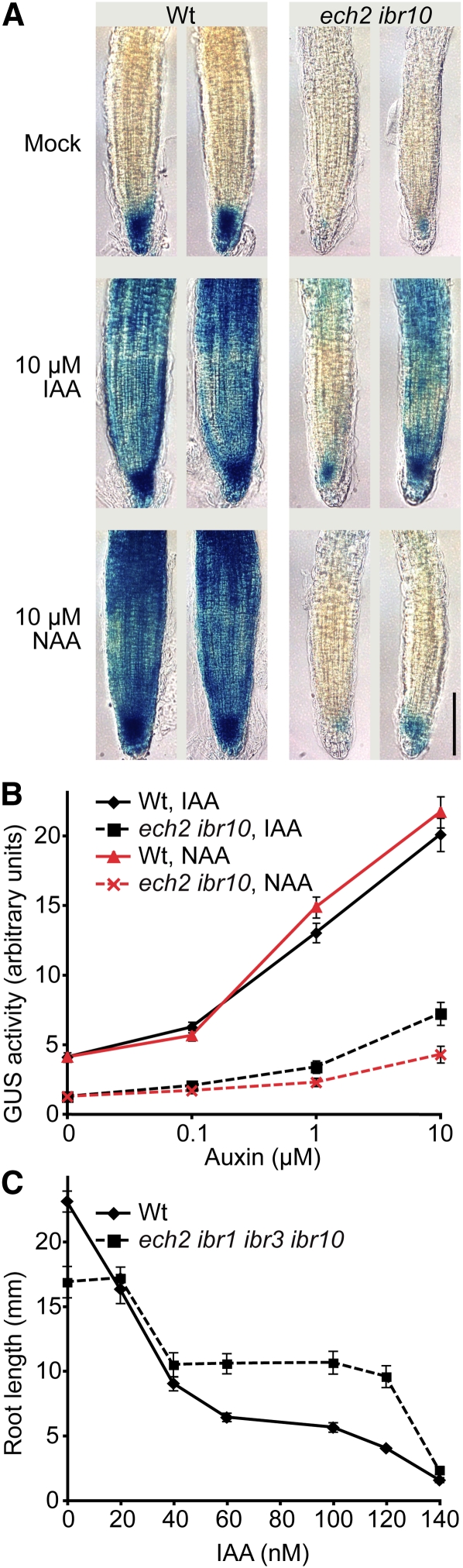
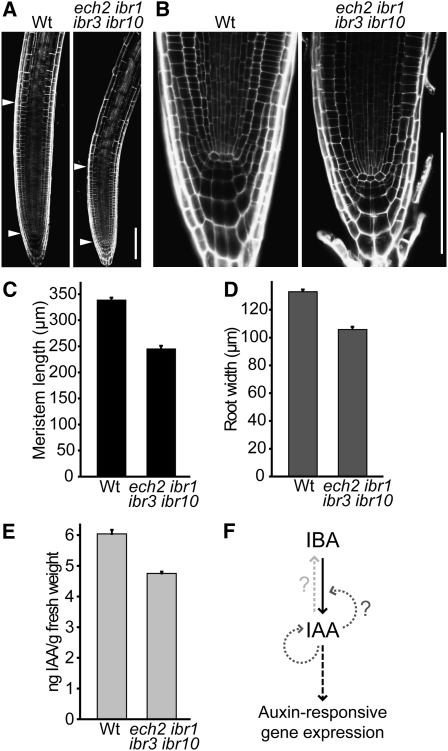
Levels of auxin, which regulates both cell division and cell elongation in plant development, are controlled by synthesis, inactivation, transport, and the use of storage forms. However, the specific contributions of various inputs to the active auxin pool are not well understood. One auxin precursor is indole-3-butyric acid (IBA), which undergoes peroxisomal β-oxidation to release free indole-3-acetic acid (IAA). We identified ENOYL-COA HYDRATASE2 (ECH2) as an enzyme required for IBA response. Combining the ech2 mutant with previously identified iba response mutants resulted in enhanced IBA resistance, diverse auxin-related developmental defects, decreased auxin-responsive reporter activity in both untreated and auxin-treated seedlings, and decreased free IAA levels. The decreased auxin levels and responsiveness, along with the associated developmental defects, uncover previously unappreciated roles for IBA-derived IAA during seedling development, establish IBA as an important auxin precursor, and suggest that IBA-to-IAA conversion contributes to the positive feedback that maintains root auxin levels.
Figures
References
-
- Adham A.R., Zolman B.K., Millius A., Bartel B. (2005). Mutations in Arabidopsis acyl-CoA oxidase genes reveal distinct and overlapping roles in β-oxidation. Plant J. 41: 859–874 - PubMed
-
- Barkawi L.S., Tam Y.Y., Tillman J.A., Normanly J., Cohen J.D. (2010). A high-throughput method for the quantitative analysis of auxins. Nat. Protoc. 5: 1609–1618 - PubMed
-
- Barkawi L.S., Tam Y.Y., Tillman J.A., Pederson B., Calio J., Al-Amier H., Emerick M., Normanly J., Cohen J.D. (2008). A high-throughput method for the quantitative analysis of indole-3-acetic acid and other auxins from plant tissue. Anal. Biochem. 372: 177–188 - PubMed
-
- Casimiro I., Beeckman T., Graham N., Bhalerao R., Zhang H., Casero P., Sandberg G., Bennett M.J. (2003). Dissecting Arabidopsis lateral root development. Trends Plant Sci. 8: 165–171 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
