

Impact of myocyte strain on cardiac myofilament activation
- PMID: 21409385
- PMCID: PMC3115504
- DOI: 10.1007/s00424-011-0952-3
Impact of myocyte strain on cardiac myofilament activation
Abstract
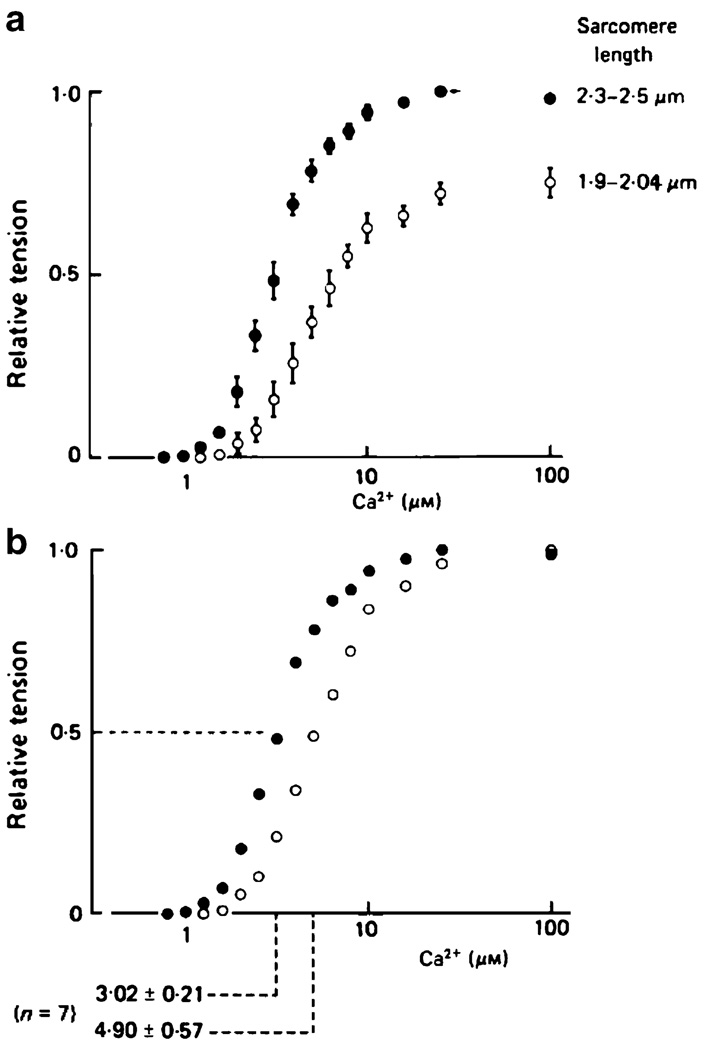
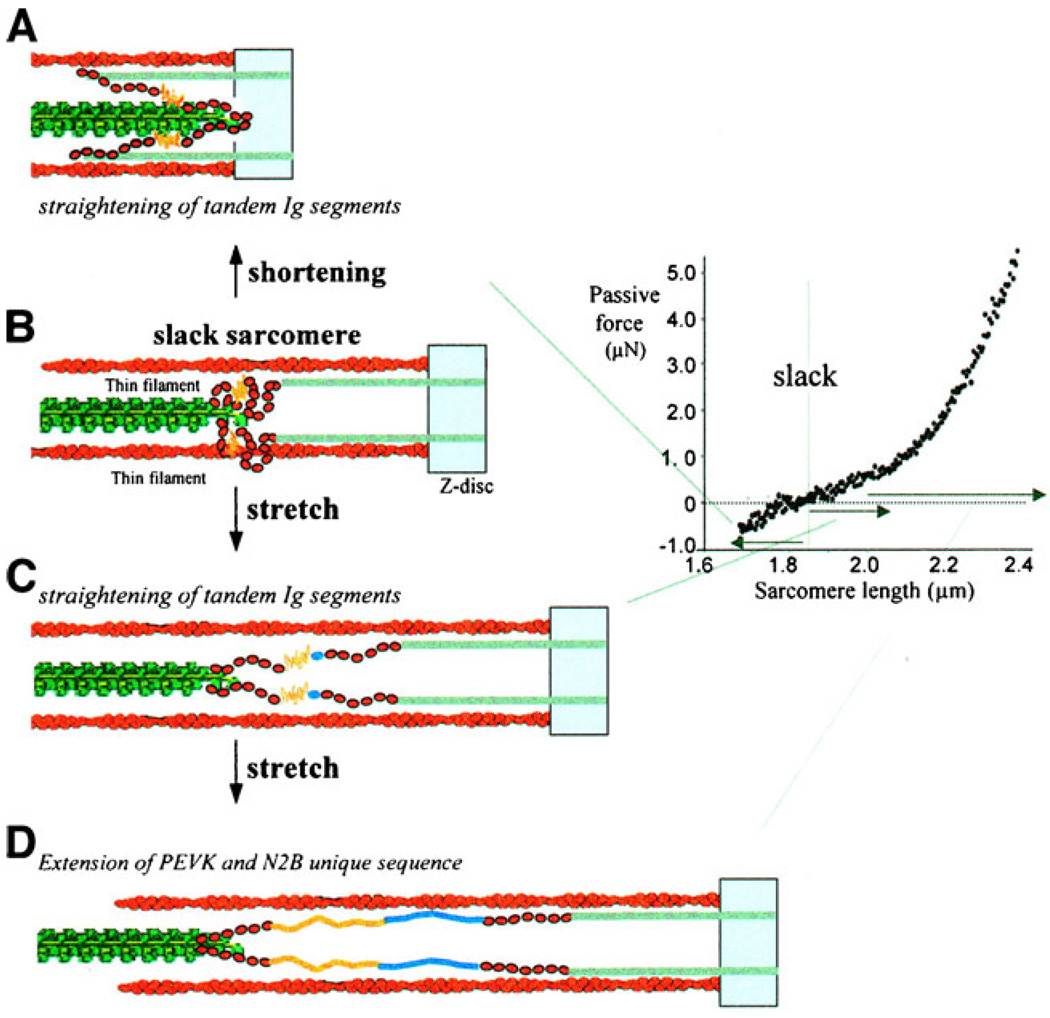
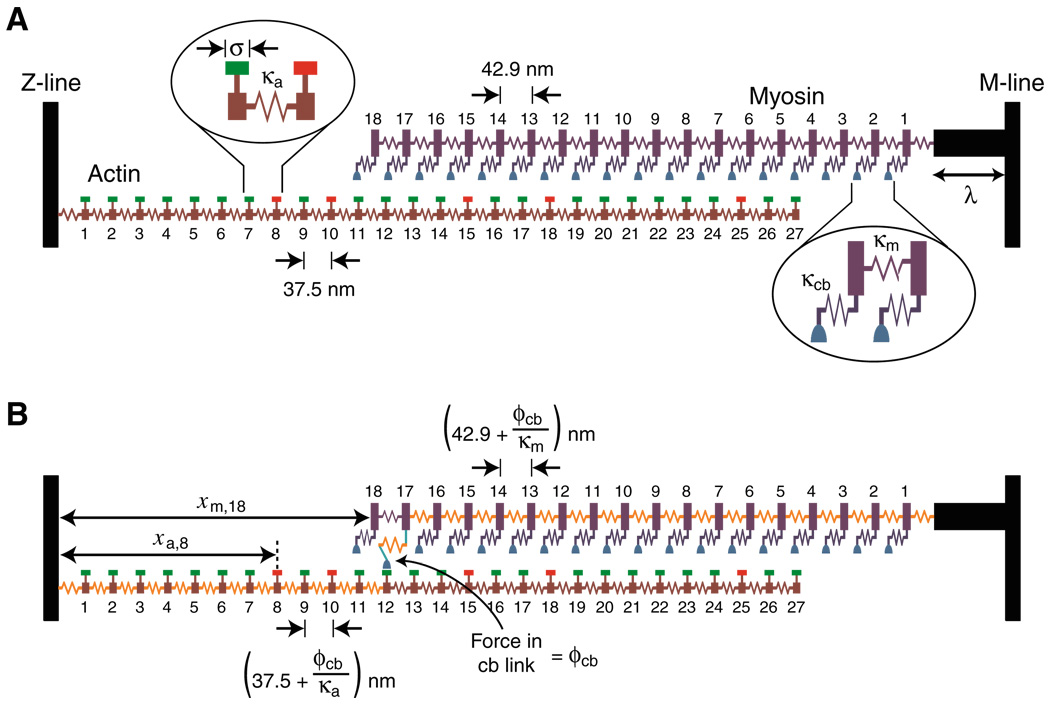
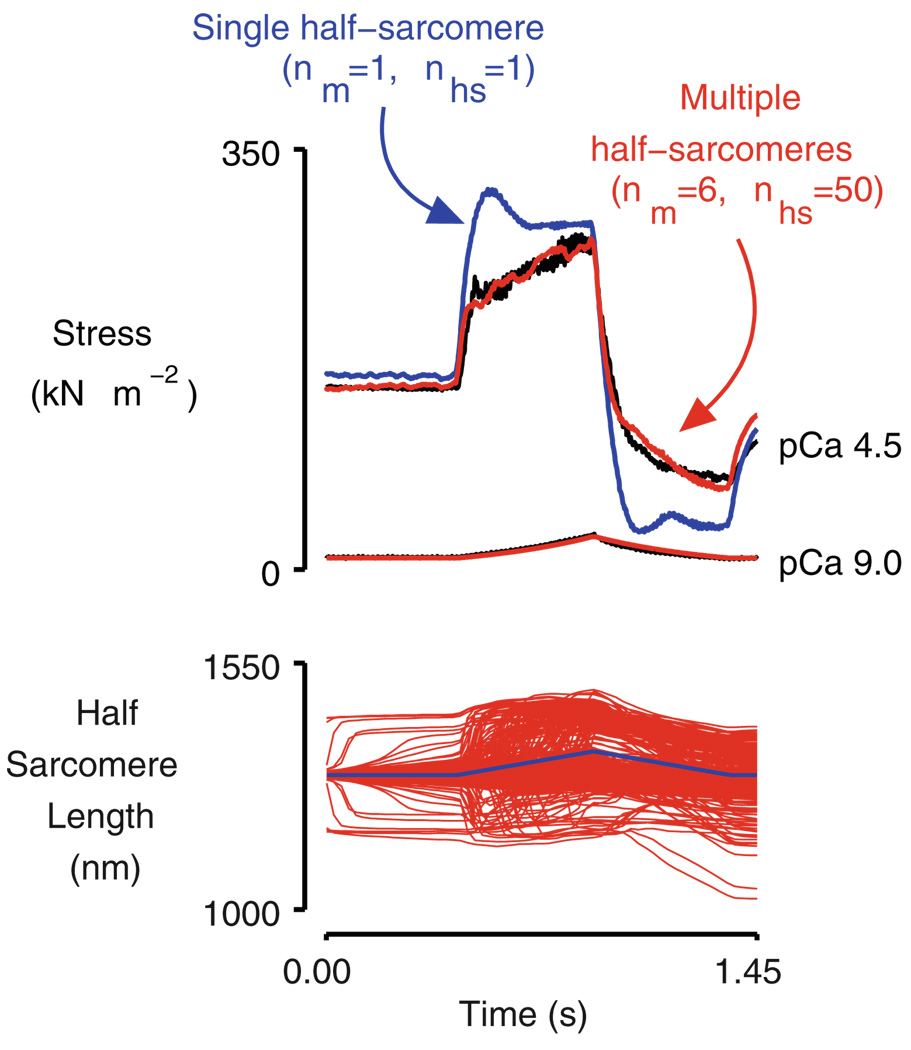
When cardiac myocytes are stretched by a longitudinal strain, they develop proportionally more active force at a given sub-maximal Ca(2+) concentration than they did at the shorter length. This is known as length-dependent activation. It is one of the most important contributors to the Frank-Starling relationship, a critical part of normal cardiovascular function. Despite intense research efforts, the mechanistic basis of the Frank-Starling relationship remains unclear. Potential mechanisms involving myofibrillar lattice spacing, titin-based effects, and cooperative activation have all been proposed. This review summarizes some of these mechanisms and discusses two additional potential theories that reflect the effects of localized strains that occur within and between half-sarcomeres. The main conclusion is that the Frank-Starling relationship is probably the integrated result of many interacting molecular mechanisms. Multiscale computational modeling may therefore provide the best way of determining the key processes that underlie length-dependent activation and their relative strengths.
Figures
Similar articles
-
Titin determines the Frank-Starling relation in early diastole.J Gen Physiol. 2003 Feb;121(2):97-110. doi: 10.1085/jgp.20028652. J Gen Physiol. 2003. PMID: 12566538 Free PMC article.
-
Titin-based modulation of calcium sensitivity of active tension in mouse skinned cardiac myocytes.Circ Res. 2001 May 25;88(10):1028-35. doi: 10.1161/hh1001.090876. Circ Res. 2001. PMID: 11375272
-
Mouse intact cardiac myocyte mechanics: cross-bridge and titin-based stress in unactivated cells.J Gen Physiol. 2011 Jan;137(1):81-91. doi: 10.1085/jgp.201010499. J Gen Physiol. 2011. PMID: 21187335 Free PMC article.
-
Titin/connectin-based modulation of the Frank-Starling mechanism of the heart.J Muscle Res Cell Motil. 2005;26(6-8):319-23. doi: 10.1007/s10974-005-9038-1. J Muscle Res Cell Motil. 2005. PMID: 16453158 Review.
-
Regional variation in myofilament length-dependent activation.Pflugers Arch. 2011 Jul;462(1):15-28. doi: 10.1007/s00424-011-0933-6. Epub 2011 Feb 19. Pflugers Arch. 2011. PMID: 21336586 Review.
Cited by
-
An analysis of deformation-dependent electromechanical coupling in the mouse heart.J Physiol. 2012 Sep 15;590(18):4553-69. doi: 10.1113/jphysiol.2012.231928. Epub 2012 May 21. J Physiol. 2012. PMID: 22615436 Free PMC article.
-
Role of the C-terminus mobile domain of cardiac troponin I in the regulation of thin filament activation in skinned papillary muscle strips.Arch Biochem Biophys. 2018 Jun 15;648:27-35. doi: 10.1016/j.abb.2018.04.014. Epub 2018 Apr 25. Arch Biochem Biophys. 2018. PMID: 29704484 Free PMC article.
-
Multiscale Modeling of Cardiovascular Function Predicts That the End-Systolic Pressure Volume Relationship Can Be Targeted via Multiple Therapeutic Strategies.Front Physiol. 2020 Aug 19;11:1043. doi: 10.3389/fphys.2020.01043. eCollection 2020. Front Physiol. 2020. PMID: 32973561 Free PMC article.
-
Length-dependent effects on cardiac contractile dynamics are different in cardiac muscle containing α- or β-myosin heavy chain.Arch Biochem Biophys. 2013 Jul 1;535(1):3-13. doi: 10.1016/j.abb.2012.10.011. Epub 2012 Oct 27. Arch Biochem Biophys. 2013. PMID: 23111184 Free PMC article.
-
Titin (TTN): from molecule to modifications, mechanics, and medical significance.Cardiovasc Res. 2022 Nov 10;118(14):2903-2918. doi: 10.1093/cvr/cvab328. Cardiovasc Res. 2022. PMID: 34662387 Free PMC article. Review.
References
-
- Allingham JS, Smith R, Rayment I. The structural basis of blebbistatin inhibition and specificity for myosin II. Nat Struct Mol Biol. 2005;12:378–379. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
