

Pattern, growth, and control
- PMID: 21414486
- PMCID: PMC3128888
- DOI: 10.1016/j.cell.2011.03.009
Pattern, growth, and control
Abstract
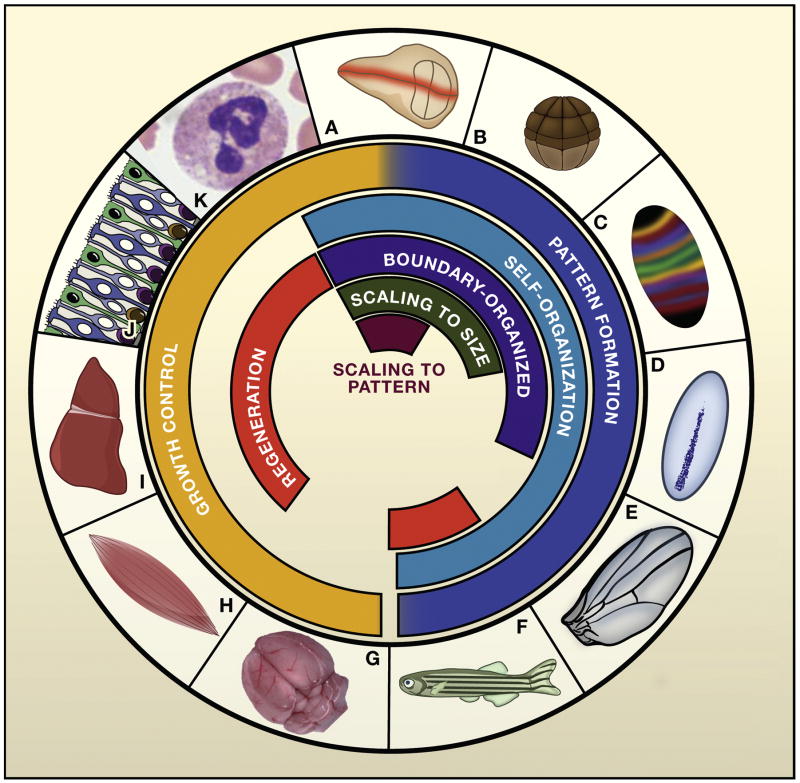
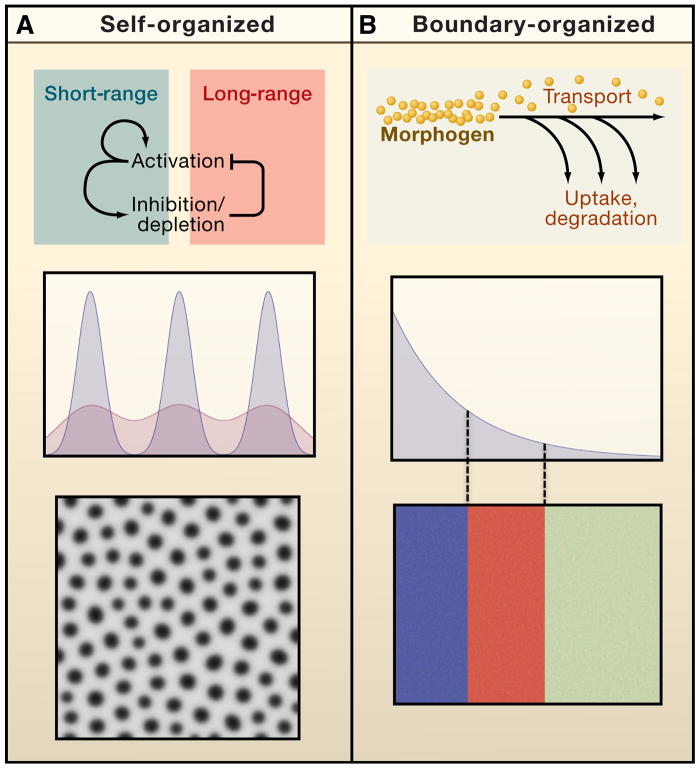
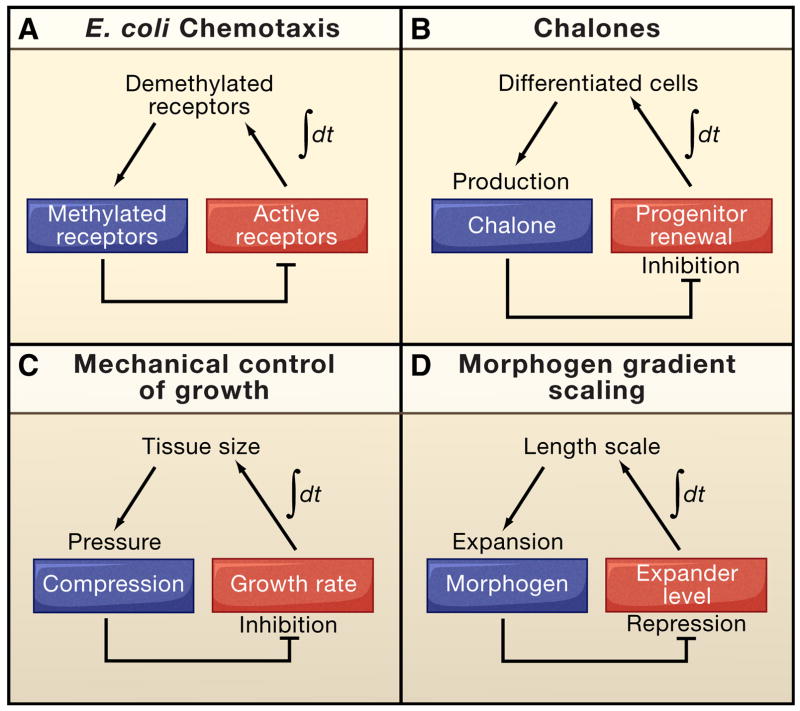
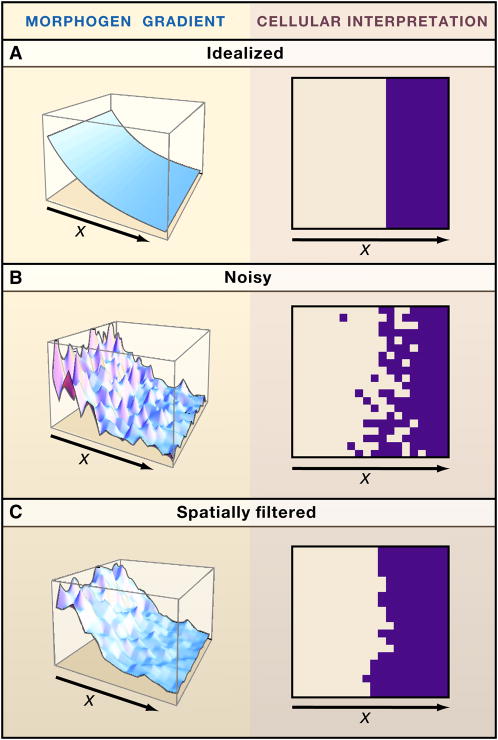
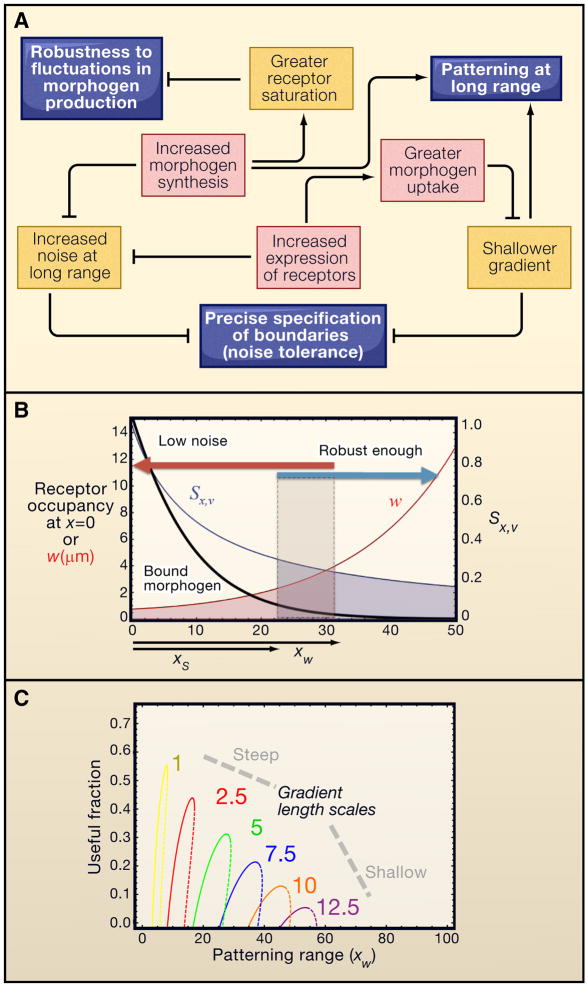
Systems biology seeks not only to discover the machinery of life but to understand how such machinery is used for control, i.e., for regulation that achieves or maintains a desired, useful end. This sort of goal-directed, engineering-centered approach also has deep historical roots in developmental biology. Not surprisingly, developmental biology is currently enjoying an influx of ideas and methods from systems biology. This Review highlights current efforts to elucidate design principles underlying the engineering objectives of robustness, precision, and scaling as they relate to the developmental control of growth and pattern formation. Examples from vertebrate and invertebrate development are used to illustrate general lessons, including the value of integral feedback in achieving set-point control; the usefulness of self-organizing behavior; the importance of recognizing and appropriately handling noise; and the absence of "free lunch." By illuminating such principles, systems biology is helping to create a functional framework within which to make sense of the mechanistic complexity of organismal development.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Aegerter-Wilmsen T, Aegerter CM, Hafen E, Basler K. Model for the regulation of size in the wing imaginal disc of Drosophila. Mech Dev. 2007;124:318–326. - PubMed
-
- Aegerter-Wilmsen T, Smith AC, Christen AJ, Aegerter CM, Hafen E, Basler K. Exploring the effects of mechanical feedback on epithelial topology. Development. 2010;137:499–506. - PubMed
-
- Alon U, Surette MG, Barkai N, Leibler S. Robustness in bacterial chemotaxis. Nature. 1999;397:168–171. - PubMed
-
- Aquino G, Endres RG. Increased accuracy of ligand sensing by receptor internalization. Phys Rev E Stat Nonlin Soft Matter Phys. 2010;81:021909. - PubMed
-
- Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: cell fate control and signal integration in development. Science. 1999;284:770–776. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
