

Theta phase precession in rat ventral striatum links place and reward information
- PMID: 21414906
- PMCID: PMC3758553
- DOI: 10.1523/JNEUROSCI.4869-10.2011
Theta phase precession in rat ventral striatum links place and reward information
Abstract
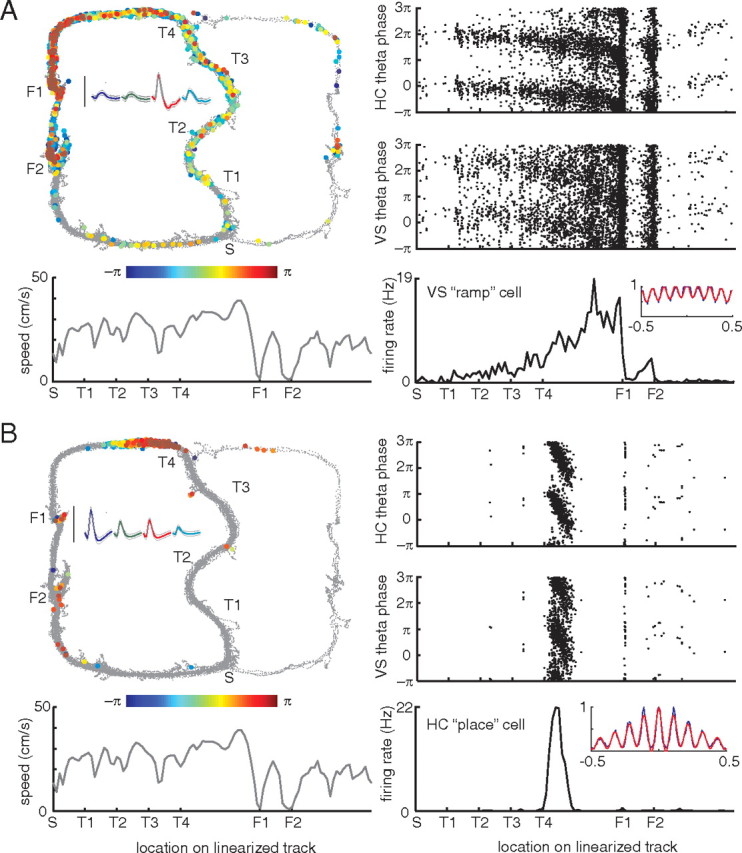
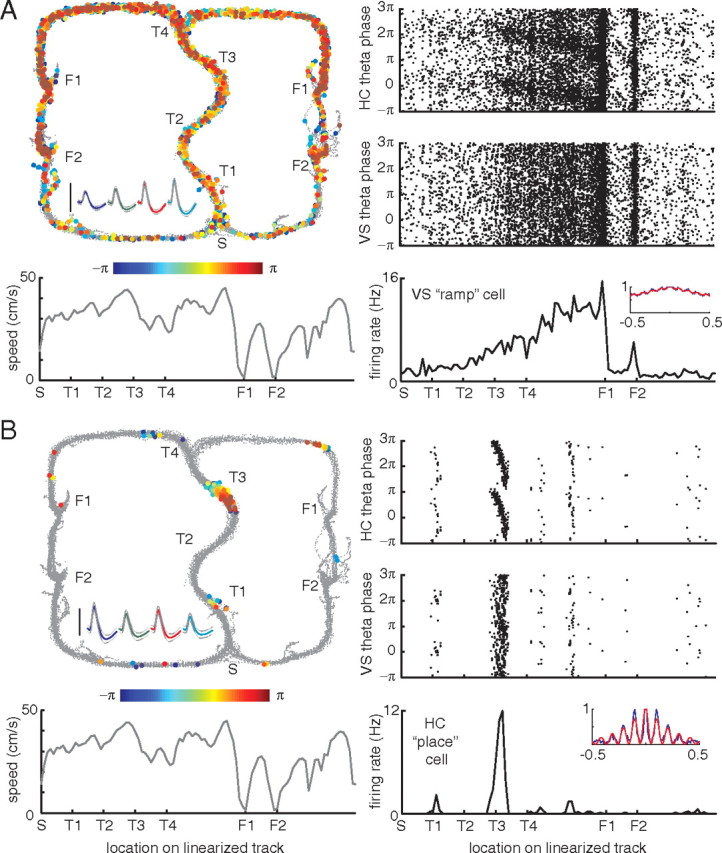
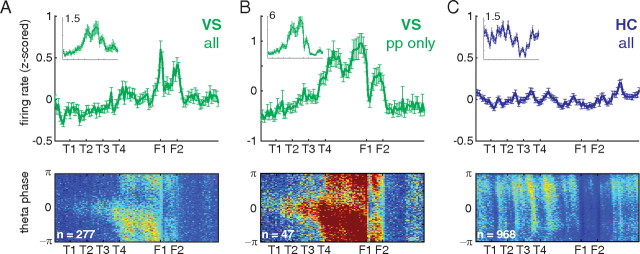
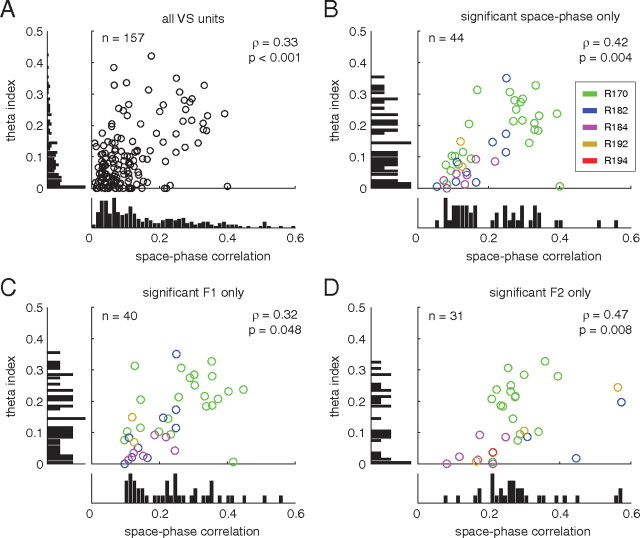
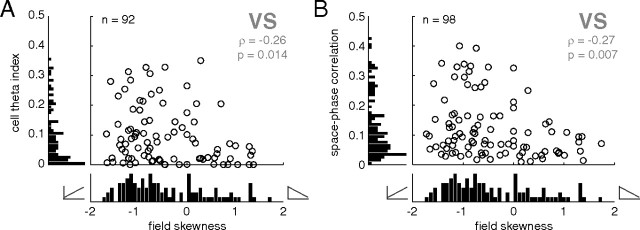
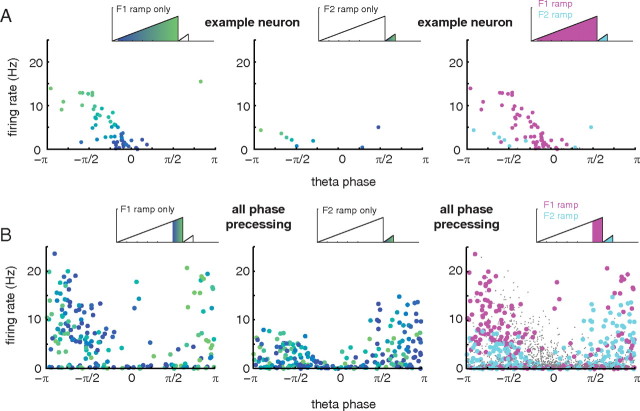
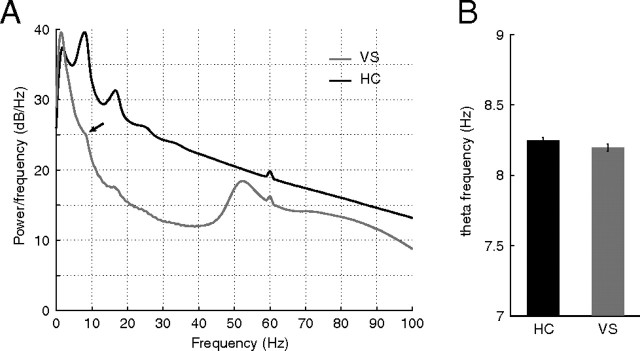
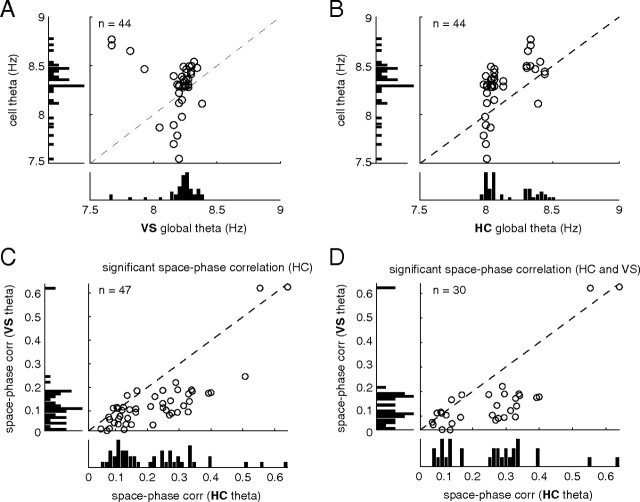
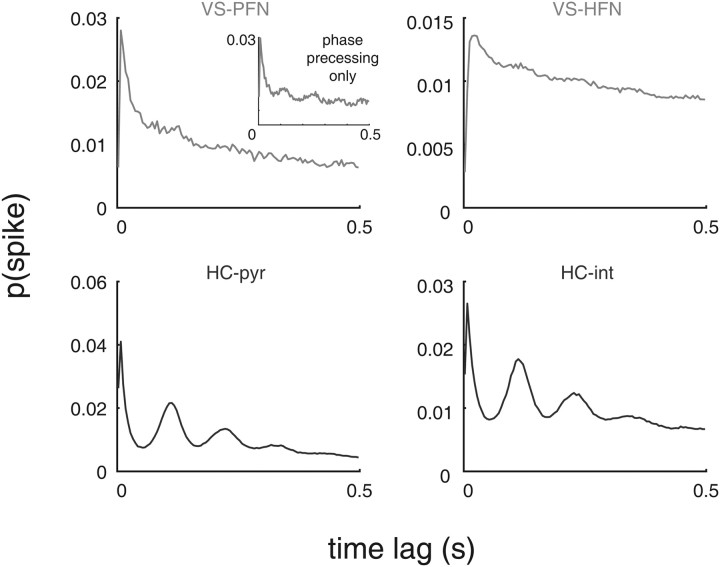
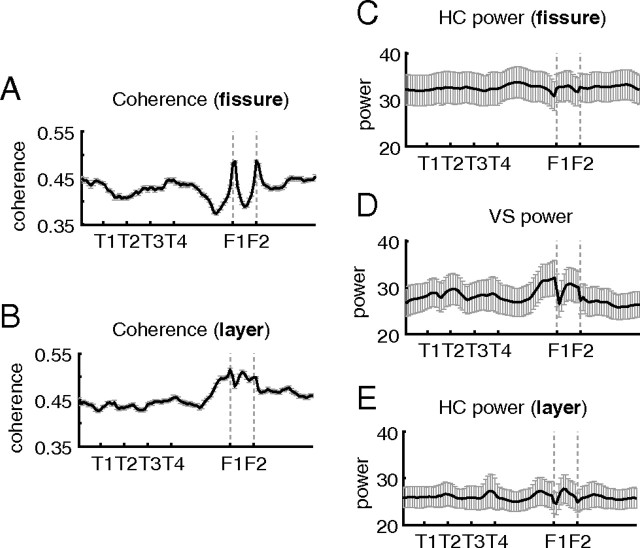
A functional interaction between the hippocampal formation and the ventral striatum is thought to contribute to the learning and expression of associations between places and rewards. However, the mechanism of how such associations may be learned and used is currently unknown. We recorded neural ensembles and local field potentials from the ventral striatum and CA1 simultaneously as rats ran a modified T-maze. Theta-modulated cells in ventral striatum almost invariably showed firing phase precession relative to the hippocampal theta rhythm. Across the population of ventral striatal cells, phase precession was preferentially associated with an anticipatory ramping of activity up to the reward sites. In contrast, CA1 population activity and phase precession were distributed more uniformly. Ventral striatal phase precession was stronger to hippocampal than ventral striatal theta and was accompanied by increased theta coherence with hippocampus, suggesting that this effect is hippocampally derived. These results suggest that the firing phase of ventral striatal neurons contains motivationally relevant information and that phase precession serves to bind hippocampal place representations to ventral striatal representations of reward.
Figures
References
-
- Atallah HE, Lopez-Paniagua D, Rudy JW, O'Reilly RC. Separate neural substrates for skill learning and performance in the ventral and dorsal striatum. Nat Neurosci. 2007;10:126–131. - PubMed
-
- Bast T, Feldon J. Hippocampal modulation of sensorimotor processes. Prog Neurobiol. 2003;70:319–345. - PubMed
-
- Benchenane K, Peyrache A, Khamassi M, Tierney PL, Gioanni Y, Battaglia FP, Wiener SI. Coherent theta oscillations and reorganization of spike timing in the hippocampal-prefrontal network upon learning. Neuron. 2010;66:921–936. - PubMed
-
- Berke JD, Okatan M, Skurski J, Eichenbaum HB. Oscillatory entrainment of striatal neurons in freely moving rats. Neuron. 2004;43:883–896. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous