

Specific inhibition of NF-Y subunits triggers different cell proliferation defects
- PMID: 21415014
- PMCID: PMC3141247
- DOI: 10.1093/nar/gkr128
Specific inhibition of NF-Y subunits triggers different cell proliferation defects
Abstract
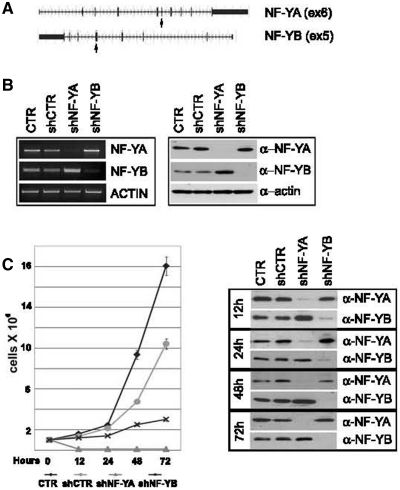
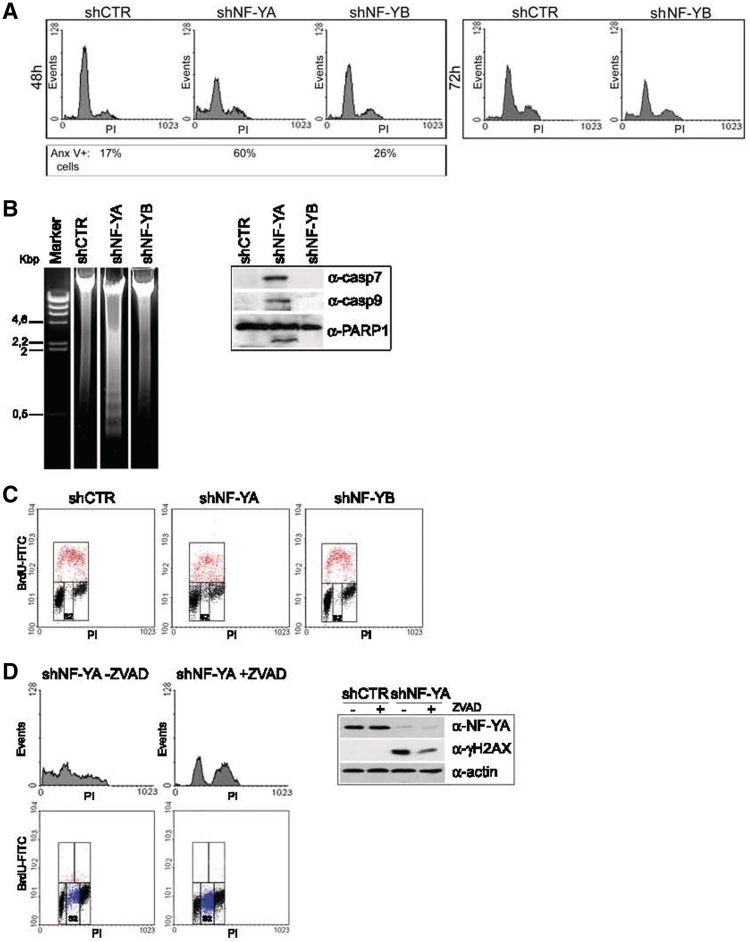
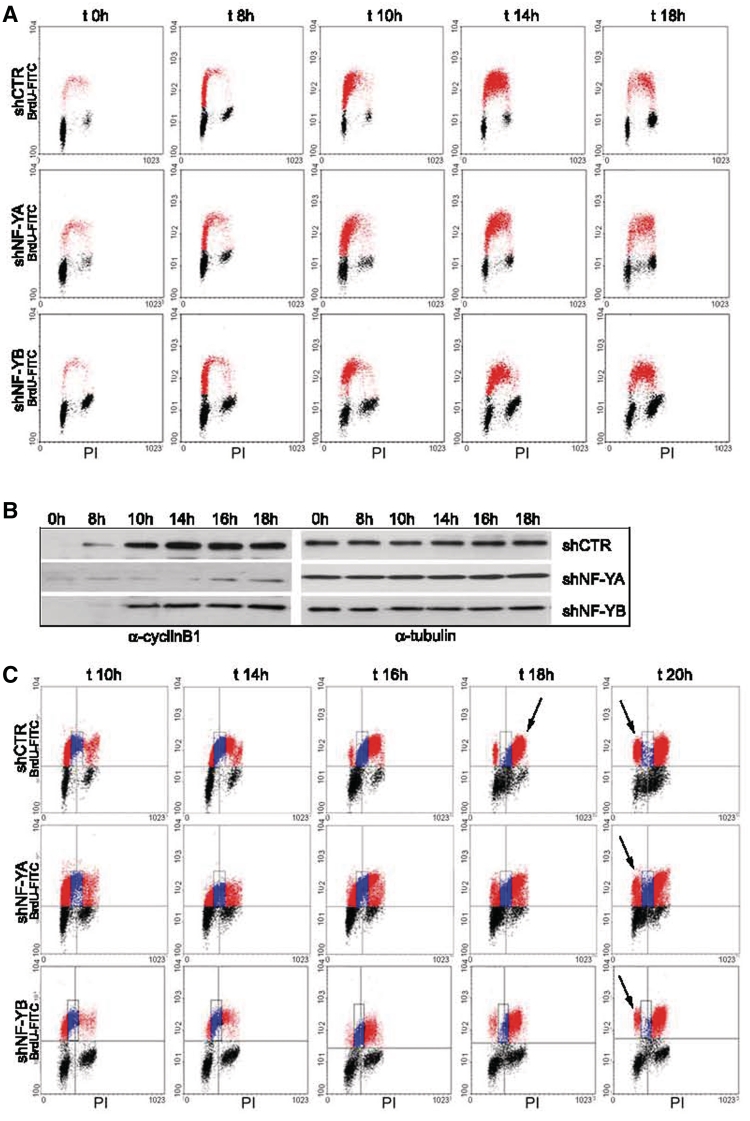
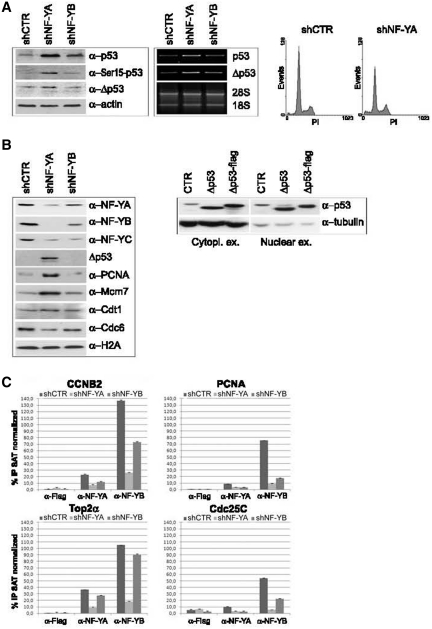
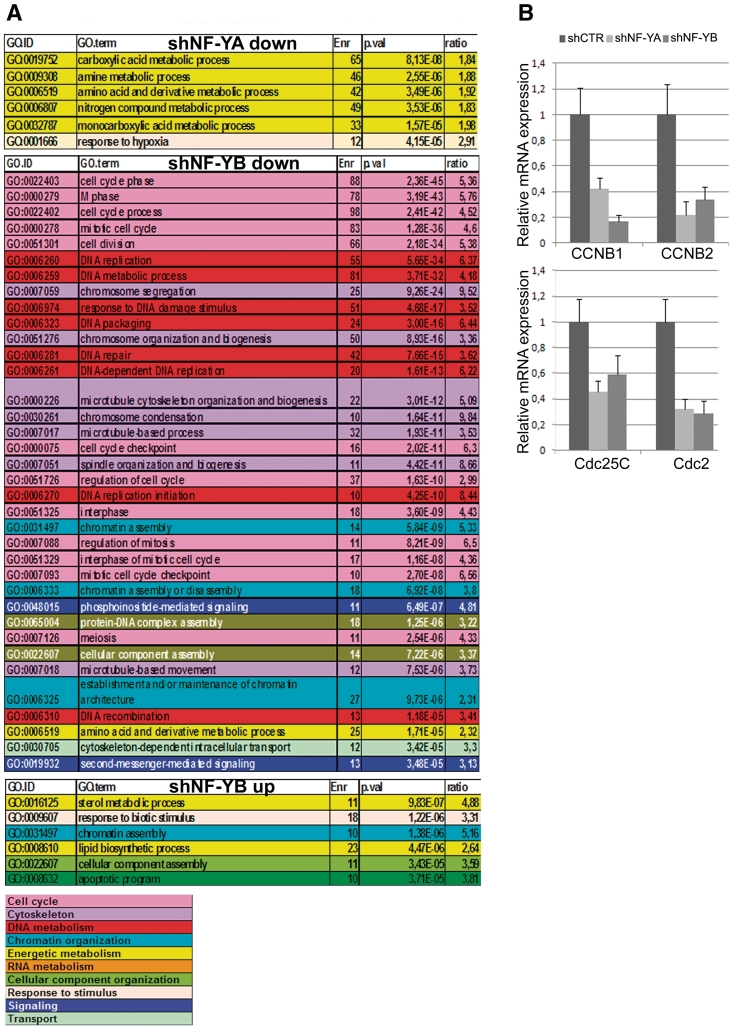
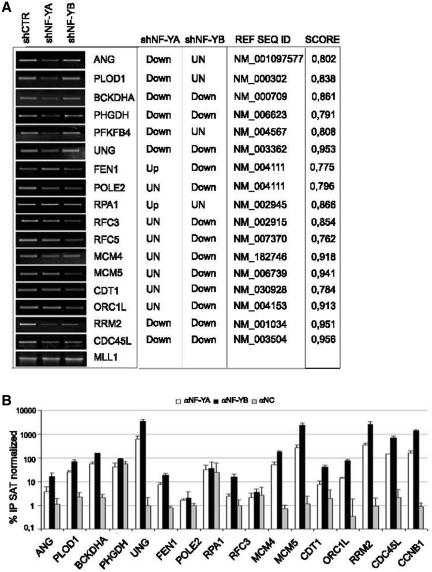
Regulated gene expression is essential for a proper progression through the cell cycle. The transcription factor NF-Y has a fundamental function in transcriptional regulation of cell cycle genes, particularly of G2/M genes. In order to investigate common and distinct functions of NF-Y subunits in cell cycle regulation, NF-YA, NF-YB and NF-YC have been silenced by shRNAs in HCT116 cells. NF-YA loss led to a delay in S-phase progression, DNA damage and apoptosis: we showed the activation of the replication checkpoint, through the recruitment of Δp53 and of the replication proteins PCNA and Mcm7 to chromatin. Differently, NF-YB depletion impaired cells from exiting G2/M, but did not interfere with S-phase progression. Gene expression analysis of NF-YA and NF-YB inactivated cells highlighted a common set of hit genes, as well as a plethora of uncommon genes, unveiling a different effect of NF-Y subunits loss on NF-Y binding to its target genes. Chromatin extracts and ChIP analysis showed that NF-YA depletion was more effective than NF-YB in hitting NF-Y recruitment to CCAAT-promoters. Our data suggest a critical role of NF-Y expression, highlighting that the lack of the single subunits are differently perceived by the cells, which activate diverse cell cycle blocks and signaling pathways.
Figures
References
-
- Mantovani R. The molecular biology of the CCAAT-binding factor NF-Y. Gene. 1999;239:15–27. - PubMed
-
- Bolognese F, Wasner M, Dohna CL, Gurtner A, Ronchi A, Muller H, Manni I, Mossner J, Piaggio G, Mantovani R, et al. The cyclin B2 promoter depends on NF-Y, a trimer whose CCAAT-binding activity is cell-cycle regulated. Oncogene. 1999;18:1845–1853. - PubMed
-
- Chang ZF, Huang DY. Regulation of thymidine kinase expression during cellular senescence. J. Biomed. Sci. 2001;8:176–183. - PubMed
-
- Marziali G, Perrotti E, Ilari R, Testa U, Coccia EM, Battistini A. Transcriptional regulation of the ferritin heavy-chain gene: the activity of the CCAAT binding factor NF-Y is modulated in heme-treated Friend leukemia cells and during monocyte-to-macrophage differentiation. Mol. Cell Biol. 1997;17:1387–1395. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
