

Roles of the spiA gene from Salmonella enteritidis in biofilm formation and virulence
- PMID: 21415117
- PMCID: PMC3167914
- DOI: 10.1099/mic.0.046185-0
Roles of the spiA gene from Salmonella enteritidis in biofilm formation and virulence
Abstract
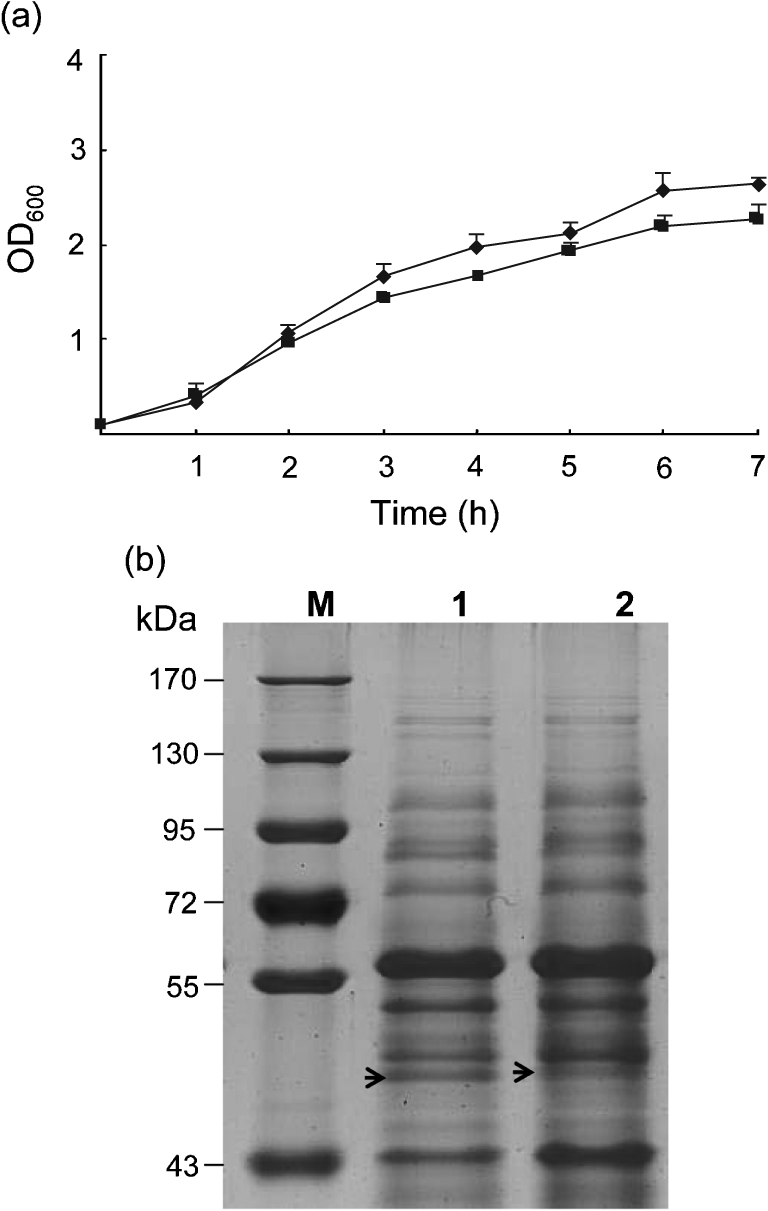
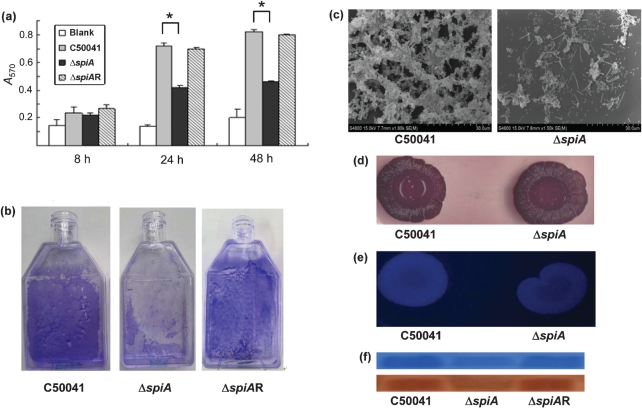
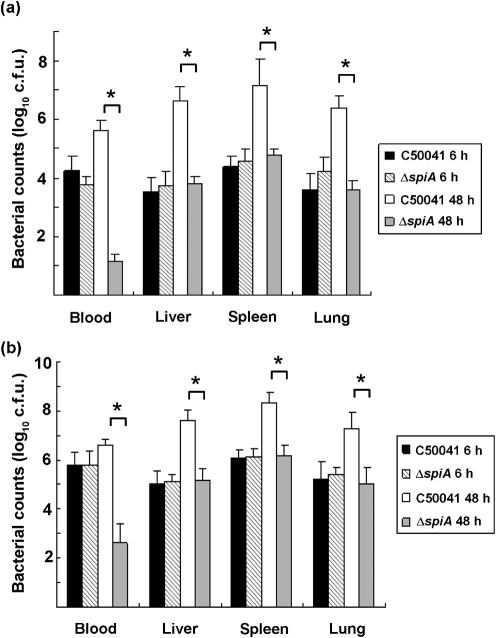
Salmonella enteritidis has emerged as one of the most important food-borne pathogens for humans, and the formation of biofilms by this species may improve its resistance to disadvantageous conditions. The spiA gene of Salmonella typhimurium is essential for its virulence in host cells. However, the roles of the spiA gene in biofilm formation and virulence of S. enteritidis remain unclear. In this study we constructed a spiA gene mutant with a suicide plasmid. Phenotypic and biological analysis revealed that the mutant was similar to the wild-type strain in growth rate, morphology, and adherence to and invasion of epithelial cells. However, the mutant showed reduced biofilm formation in a quantitative microtitre assay and by scanning electron microscopy, and significantly decreased curli production and intracellular proliferation of macrophages during the biofilm phase. In addition, the spiA mutant was attenuated in a mouse model in both the exponential growth and biofilm phases. These data indicate that the spiA gene is involved in both biofilm formation and virulence of S. enteritidis.
Figures
References
-
- Anriany Y., Sahu S. N., Wessels K. R., McCann L. M., Joseph S. W. (2006). Alteration of the rugose phenotype in waaG and ddhC mutants of Salmonella enterica serovar Typhimurium DT104 is associated with inverse production of curli and cellulose. Appl Environ Microbiol 72, 5002–5012. 10.1128/AEM.02868-05.. - DOI - PMC - PubMed
-
- Dong H., Zhang X., Pan Z., Peng D., Liu X. (2008). [Identification of genes for biofilm formation in a Salmonella enteritidis strain by transposon mutagenesis]. Wei Sheng Wu Xue Bao 48, 869–873. (in Chinese).. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources