

Dual mTORC2/mTORC1 targeting results in potent suppressive effects on acute myeloid leukemia (AML) progenitors
- PMID: 21415215
- PMCID: PMC3131493
- DOI: 10.1158/1078-0432.CCR-10-2285
Dual mTORC2/mTORC1 targeting results in potent suppressive effects on acute myeloid leukemia (AML) progenitors
Abstract
Purpose: To determine whether mTORC2 and rapamycin-insensitive (RI)-mTORC1 complexes are present in acute myeloid leukemia (AML) cells and to examine the effects of dual mTORC2/mTORC1 inhibition on primitive AML leukemic progenitors.
Experimental design: Combinations of different experimental approaches were used, including immunoblotting to detect phosphorylated/activated forms of elements of the mTOR pathway in leukemic cell lines and primary AML blasts; cell-proliferation assays; direct assessment of mRNA translation in polysomal fractions of leukemic cells; and clonogenic assays in methylcellulose to evaluate leukemic progenitor-colony formation.
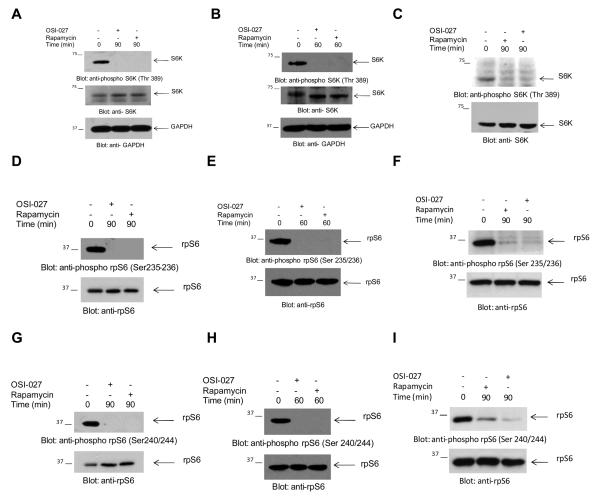
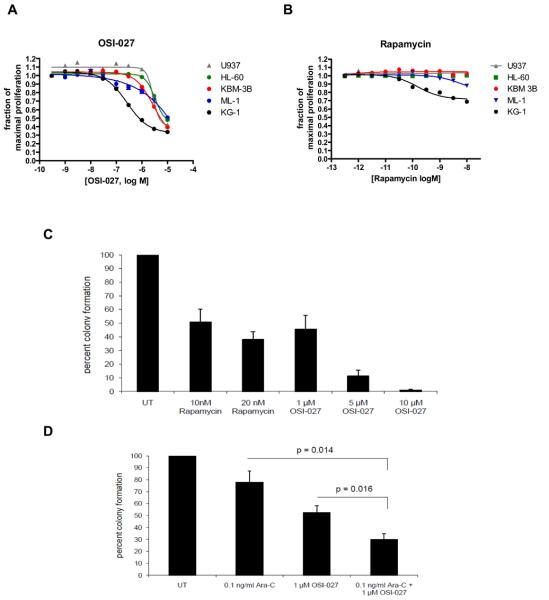
Results: mTORC2 complexes are active in AML cells and play critical roles in leukemogenesis. RI-mTORC1 complexes are also formed and regulate the activity of the translational repressor 4E-BP1 in AML cells. OSI-027 blocks mTORC1 and mTORC2 activities and suppresses mRNA translation of cyclin D1 and other genes that mediate proliferative responses in AML cells. Moreover, OSI-027 acts as a potent suppressor of primitive leukemic precursors from AML patients and is much more effective than rapamycin in eliciting antileukemic effects in vitro.
Conclusions: Dual targeting of mTORC2 and mTORC1 results in potent suppressive effects on primitive leukemic progenitors from AML patients. Inhibition of the mTOR catalytic site with OSI-027 results in suppression of both mTORC2 and RI-mTORC1 complexes and elicits much more potent antileukemic responses than selective mTORC1 targeting with rapamycin.
Figures




Similar articles
-
Autophagy is a survival mechanism of acute myelogenous leukemia precursors during dual mTORC2/mTORC1 targeting.Clin Cancer Res. 2014 May 1;20(9):2400-9. doi: 10.1158/1078-0432.CCR-13-3218. Epub 2014 Mar 7. Clin Cancer Res. 2014. PMID: 24610825 Free PMC article.
-
Preclinical characterization of OSI-027, a potent and selective inhibitor of mTORC1 and mTORC2: distinct from rapamycin.Mol Cancer Ther. 2011 Aug;10(8):1394-406. doi: 10.1158/1535-7163.MCT-10-1099. Epub 2011 Jun 14. Mol Cancer Ther. 2011. PMID: 21673091
-
Protein synthesis is resistant to rapamycin and constitutes a promising therapeutic target in acute myeloid leukemia.Blood. 2009 Aug 20;114(8):1618-27. doi: 10.1182/blood-2008-10-184515. Epub 2009 May 20. Blood. 2009. PMID: 19458359
-
Targeting translation in acute myeloid leukemia: a new paradigm for therapy?Cell Cycle. 2009 Dec;8(23):3893-9. doi: 10.4161/cc.8.23.10091. Cell Cycle. 2009. PMID: 19934662 Review.
-
Targeting the translational apparatus to improve leukemia therapy: roles of the PI3K/PTEN/Akt/mTOR pathway.Leukemia. 2011 Jul;25(7):1064-79. doi: 10.1038/leu.2011.46. Epub 2011 Mar 25. Leukemia. 2011. PMID: 21436840 Review.
Cited by
-
New strategies in acute myeloid leukemia: redefining prognostic markers to guide therapy.Clin Cancer Res. 2012 Oct 1;18(19):5163-71. doi: 10.1158/1078-0432.CCR-12-0313. Epub 2012 Aug 14. Clin Cancer Res. 2012. PMID: 22893630 Free PMC article.
-
S6K1 regulates hematopoietic stem cell self-renewal and leukemia maintenance.J Clin Invest. 2016 Jul 1;126(7):2621-5. doi: 10.1172/JCI84565. Epub 2016 Jun 13. J Clin Invest. 2016. PMID: 27294524 Free PMC article.
-
Regulation of the kinase RSK1 by arsenic trioxide and generation of antileukemic responses.Cancer Biol Ther. 2013 May;14(5):411-6. doi: 10.4161/cbt.23760. Epub 2013 Feb 1. Cancer Biol Ther. 2013. PMID: 23377826 Free PMC article.
-
AZD1208, a potent and selective pan-Pim kinase inhibitor, demonstrates efficacy in preclinical models of acute myeloid leukemia.Blood. 2014 Feb 6;123(6):905-13. doi: 10.1182/blood-2013-04-495366. Epub 2013 Dec 20. Blood. 2014. PMID: 24363397 Free PMC article.
-
Targeting mTOR for the treatment of AML. New agents and new directions.Oncotarget. 2011 Jun;2(6):510-7. doi: 10.18632/oncotarget.290. Oncotarget. 2011. PMID: 21680954 Free PMC article. Review.
References
-
- Lowenberg B, Downing JR, Burnett A. Acute myeloid leukemia. N Engl J Med. 1999;341:1051–62. - PubMed
-
- Tallman MS, Gilliland DG, Rowe JM. Drug therapy for acute myeloid leukemia. Blood. 2005;106:1154–63. - PubMed
-
- Gilliland DG, Tallman MS. Focus on acute leukemias. Cancer Cell. 2002;1:417–20. - PubMed
-
- Cammenga J. Gatekeeper pathways and cellular background in the pathogenesis and therapy of AML. Leukemia. 2005;19:1719–28. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
