

Micro-evolution of toxicant tolerance: from single genes to the genome's tangled bank
- PMID: 21416112
- PMCID: PMC3081431
- DOI: 10.1007/s10646-011-0631-3
Micro-evolution of toxicant tolerance: from single genes to the genome's tangled bank
Abstract
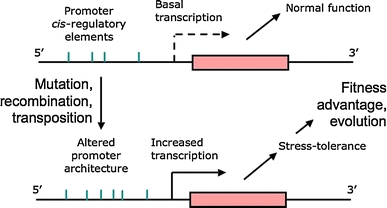
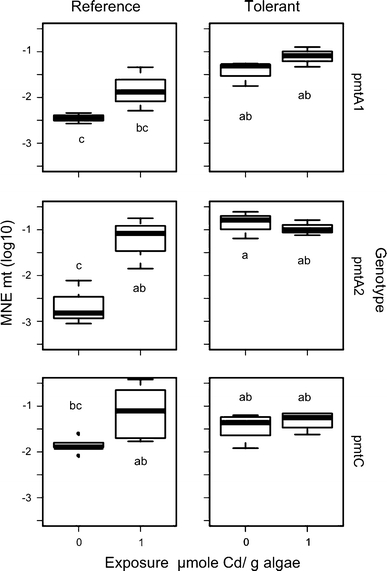
Two case-studies published 55 years ago became textbook examples of evolution in action: DDT resistance in houseflies (Busvine) and the rise of melanic forms of the peppered moth (Kettlewell). Now, many years later, molecular studies have elucidated in detail the mechanisms conferring resistance. In this paper we focus on the case of metal tolerance in a soil-living arthropod, Orchesella cincta, and provide new evidence on the transcriptional regulation of a gene involved in stress tolerance, metallothionein. Evolution of resistance is often ascribed to cis-regulatory change of such stress-combatting genes. For example, DDT resistance in the housefly is due to insertion of a mobile element into the promoter of Cyp6g1, and overexpression of this gene allows rapid metabolism of DDT. The discovery of these mechanisms has promoted the idea that resistance to environmental toxicants can be brought about by relatively simple genetic changes, involving up-regulation, duplication or structural alteration of a single-gene. Similarly, the work on O. cincta shows that populations from metal-polluted mining sites have a higher constitutive expression of the cadmium-induced metallothionein (Mt) gene. Moreover, its promoter appears to include a large degree of polymorphism; Mt promoter alleles conferring high expression in cell-based bioreporter assays were shown to occur at higher frequency in populations living at polluted sites. The case is consistent with classical examples of micro-evolution through altered cis-regulation of a key gene. However, new data on qPCR analysis of gene expression in homozygous genotypes with both reference and metal-tolerant genetic backgrounds, show that Mt expression of the same pMt homozygotes depends on the origin of the population. This suggests that trans-acting factors are also important in the regulation of Mt expression and its evolution. So the idea that metal tolerance in Orchesella can be viewed as a single-gene adaptation must be abandoned. These data, added to a genome-wide gene expression profiling study reported earlier shows that evolution of tolerance takes place in a complicated molecular network, not unlike an internal tangled bank.
© The Author(s) 2011. This article is published with open access at Springerlink.com
Figures

References
-
- Carroll SB, Grenier JK, Weatherbee SD. From DNA to diversity. Malden: Blackwell; 2005.
-
- Darwin C. The origin of species. Harmondsworth: Penguin Books Ltd; 1859.
-
- Ernsting G, Fokkema DS. Antennal damage and regeneration in springtails (Collembola) in relation to predation. Neth J Zool. 1983;33:476–484. doi: 10.1163/002829683X00200. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
