

Diversity of bacterial type II toxin-antitoxin systems: a comprehensive search and functional analysis of novel families
- PMID: 21422074
- PMCID: PMC3141249
- DOI: 10.1093/nar/gkr131
Diversity of bacterial type II toxin-antitoxin systems: a comprehensive search and functional analysis of novel families
Abstract
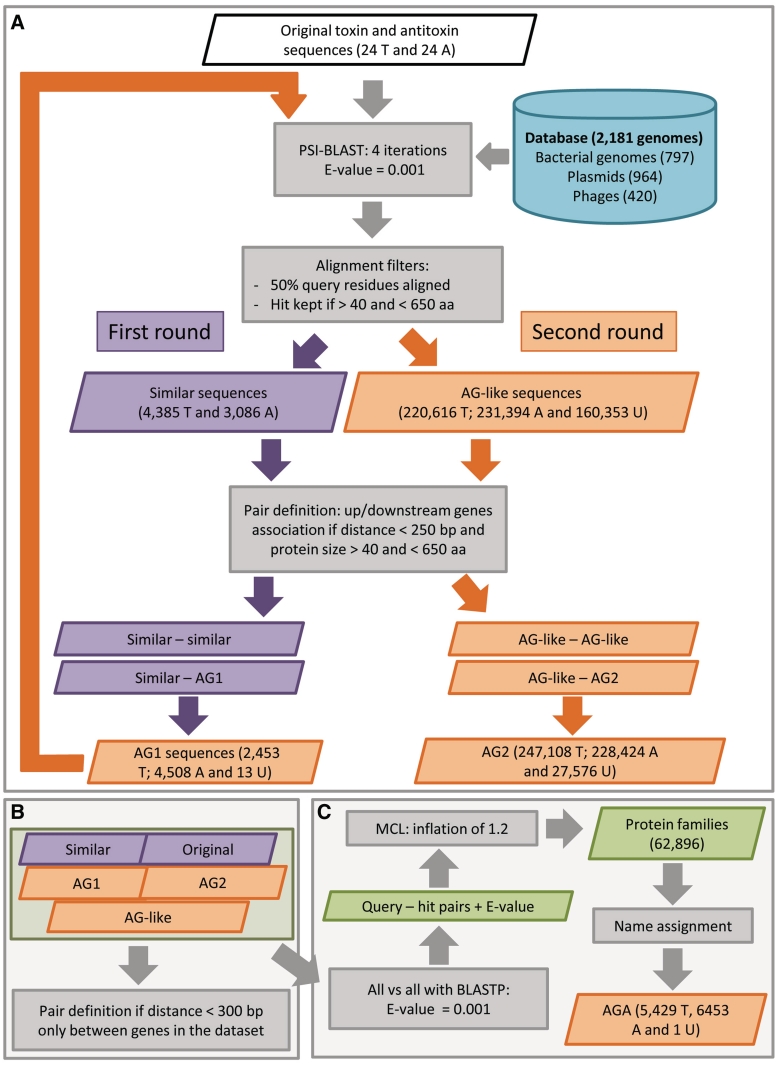
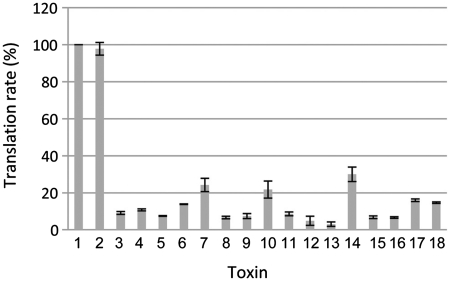
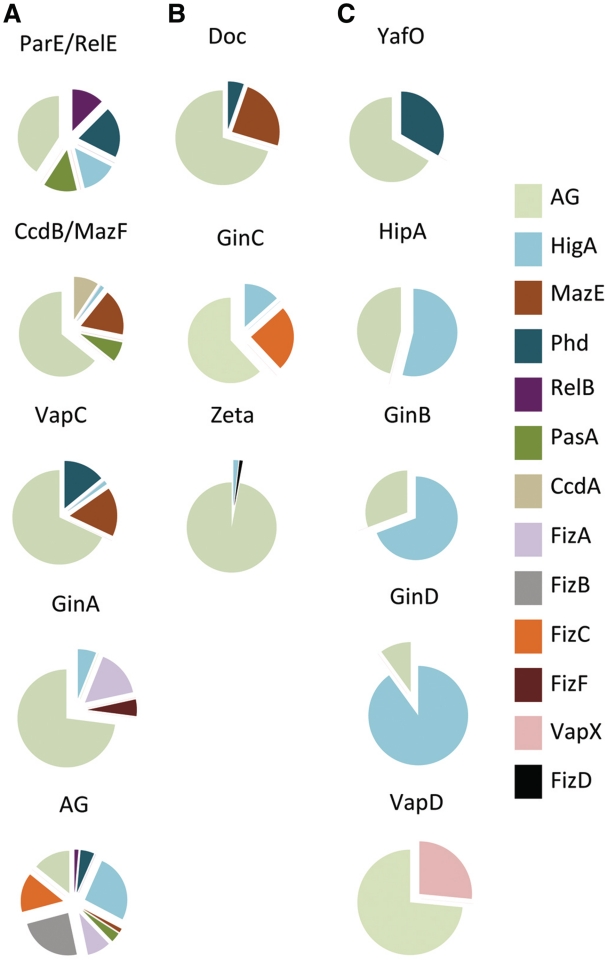
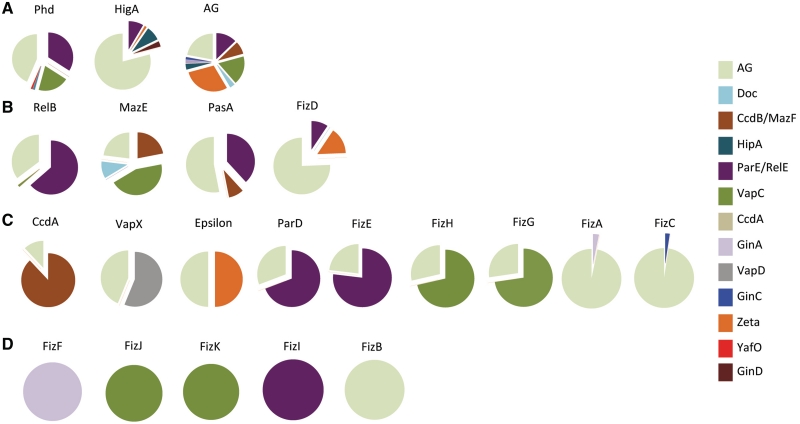
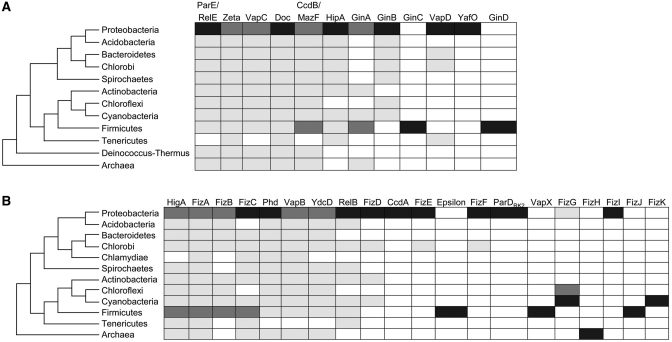
Type II toxin-antitoxin (TA) systems are generally composed of two genes organized in an operon, encoding a labile antitoxin and a stable toxin. They were first discovered on plasmids where they contribute to plasmid stability by a phenomenon denoted as 'addiction', and subsequently in bacterial chromosomes. To discover novel families of antitoxins and toxins, we developed a bioinformatics approach based on the 'guilt by association' principle. Extensive experimental validation in Escherichia coli of predicted antitoxins and toxins increased significantly the number of validated systems and defined novel toxin and antitoxin families. Our data suggest that toxin families as well as antitoxin families originate from distinct ancestors that were assembled multiple times during evolution. Toxin and antitoxin families found on plasmids tend to be promiscuous and widespread, indicating that TA systems move through horizontal gene transfer. We propose that due to their addictive properties, TA systems are likely to be maintained in chromosomes even though they do not necessarily confer an advantage to their bacterial hosts. Therefore, addiction might play a major role in the evolutionary success of TA systems both on mobile genetic elements and in bacterial chromosomes.
Figures



References
-
- Arcus VL, Rainey PB, Turner SJ. The PIN-domain toxin–antitoxin array in mycobacteria. Trends Microbiol. 2005;13:360–365. - PubMed
-
- Gerdes K, Christensen SK, Lobner-Olesen A. Prokaryotic toxin–antitoxin stress response loci. Nat. Rev. Microbiol. 2005;3:371–382. - PubMed
-
- Yarmolinsky MB. Programmed cell death in bacterial populations. Science. 1995;267:836–837. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
