

Global analysis of the relationship between JIL-1 kinase and transcription
- PMID: 21423663
- PMCID: PMC3053325
- DOI: 10.1371/journal.pgen.1001327
Global analysis of the relationship between JIL-1 kinase and transcription
Abstract
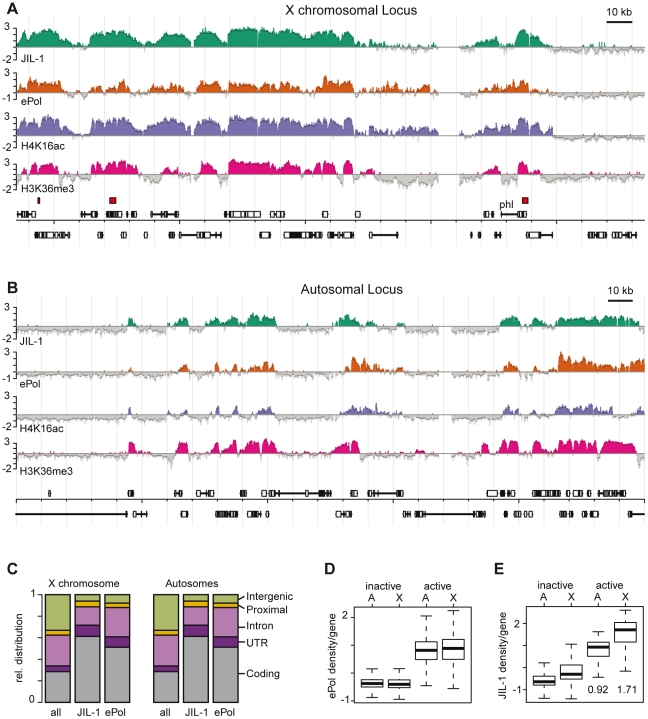
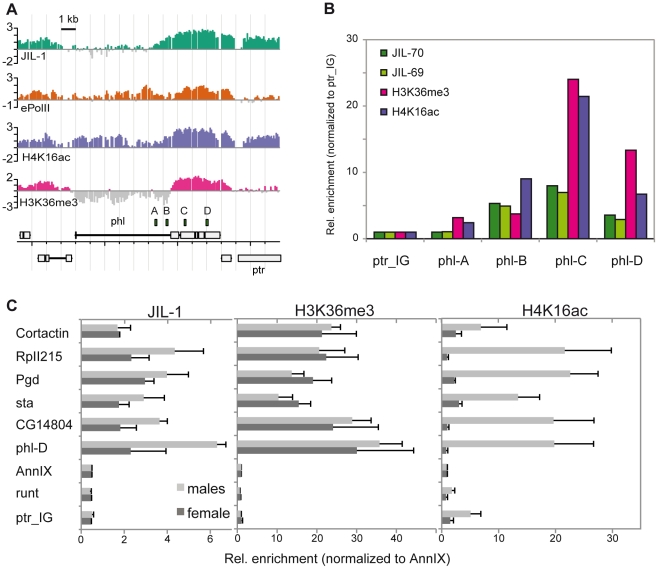
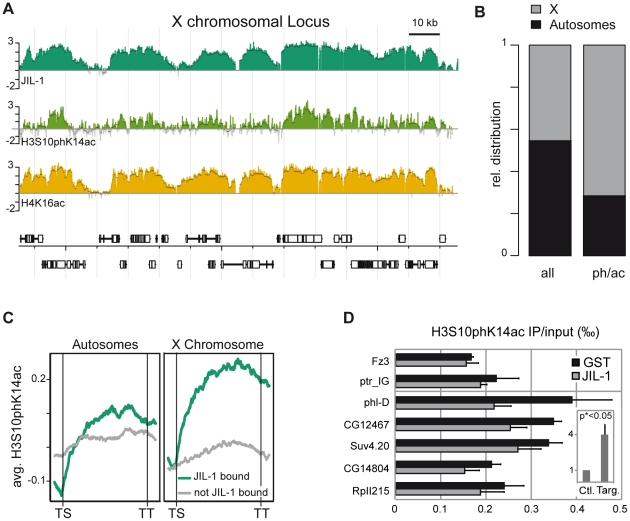
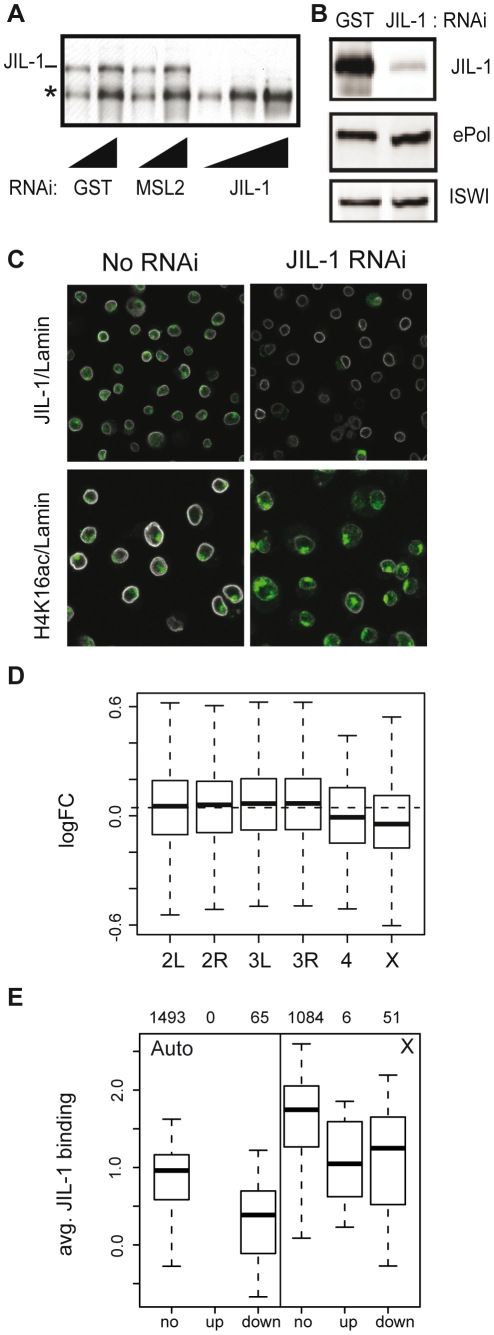
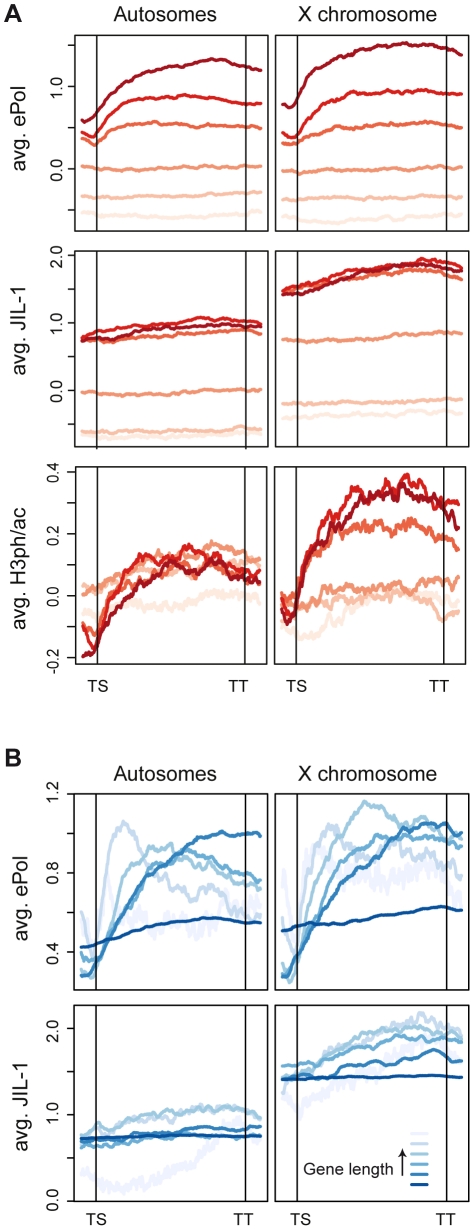
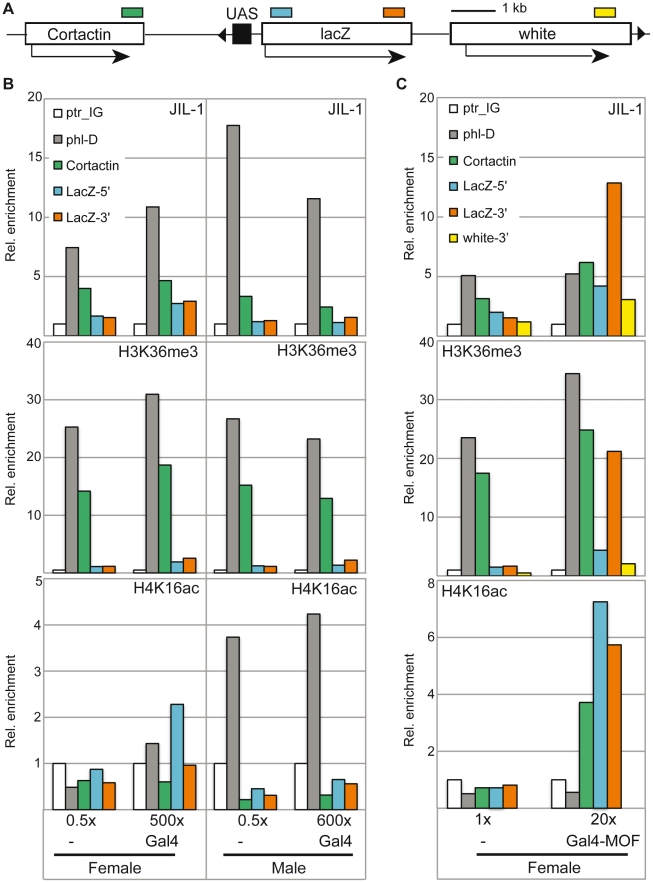
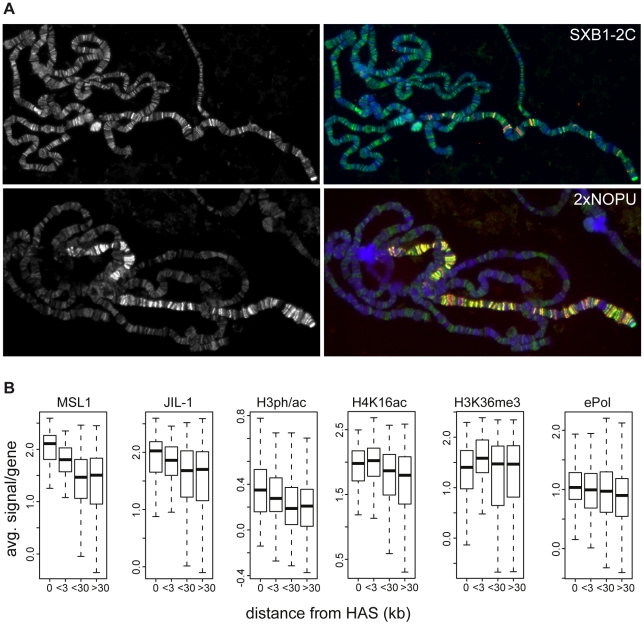
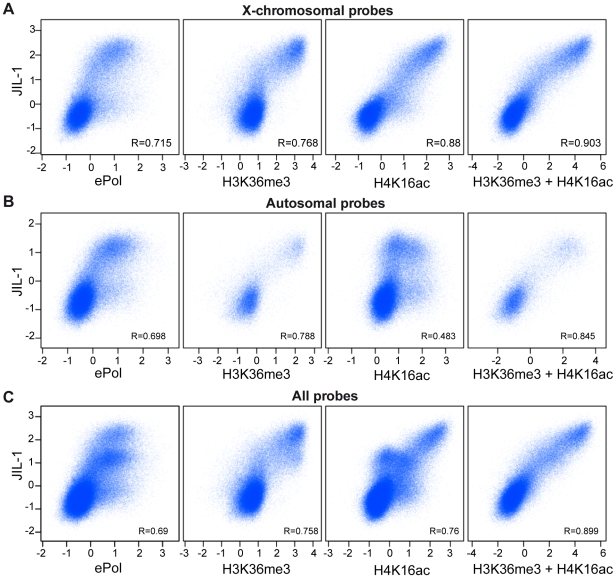
The ubiquitous tandem kinase JIL-1 is essential for Drosophila development. Its role in defining decondensed domains of larval polytene chromosomes is well established, but its involvement in transcription regulation has remained controversial. For a first comprehensive molecular characterisation of JIL-1, we generated a high-resolution, chromosome-wide interaction profile of the kinase in Drosophila cells and determined its role in transcription. JIL-1 binds active genes along their entire length. The presence of the kinase is not proportional to average transcription levels or polymerase density. Comparison of JIL-1 association with elongating RNA polymerase and a variety of histone modifications suggests two distinct targeting principles. A basal level of JIL-1 binding can be defined that correlates best with the methylation of histone H3 at lysine 36, a mark that is placed co-transcriptionally. The additional acetylation of H4K16 defines a second state characterised by approximately twofold elevated JIL-1 levels, which is particularly prominent on the dosage-compensated male X chromosome. Phosphorylation of the histone H3 N-terminus by JIL-1 in vitro is compatible with other tail modifications. In vivo, phosphorylation of H3 at serine 10, together with acetylation at lysine 14, creates a composite histone mark that is enriched at JIL-1 binding regions. Its depletion by RNA interference leads to a modest, but significant, decrease of transcription from the male X chromosome. Collectively, the results suggest that JIL-1 participates in a complex histone modification network that characterises active, decondensed chromatin. We hypothesise that one specific role of JIL-1 may be to reinforce, rather than to establish, the status of active chromatin through the phosphorylation of histone H3 at serine 10.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Domain requirements of the JIL-1 tandem kinase for histone H3 serine 10 phosphorylation and chromatin remodeling in vivo.J Biol Chem. 2013 Jul 5;288(27):19441-9. doi: 10.1074/jbc.M113.464271. Epub 2013 May 30. J Biol Chem. 2013. PMID: 23723094 Free PMC article.
-
Sequence-specific targeting of dosage compensation in Drosophila favors an active chromatin context.PLoS Genet. 2012;8(4):e1002646. doi: 10.1371/journal.pgen.1002646. Epub 2012 Apr 26. PLoS Genet. 2012. PMID: 22570616 Free PMC article.
-
GAL4 induces transcriptionally active puff in the absence of dSAGA- and ATAC-specific chromatin acetylation in the Drosophila melanogaster polytene chromosome.Chromosoma. 2009 Aug;118(4):513-26. doi: 10.1007/s00412-009-0215-7. Epub 2009 May 2. Chromosoma. 2009. PMID: 19412618
-
Dosage Compensation of the X Chromosome: A Complex Epigenetic Assignment Involving Chromatin Regulators and Long Noncoding RNAs.Annu Rev Biochem. 2018 Jun 20;87:323-350. doi: 10.1146/annurev-biochem-062917-011816. Epub 2018 Apr 18. Annu Rev Biochem. 2018. PMID: 29668306 Review.
-
The structure-function link of compensated chromatin in Drosophila.Curr Opin Genet Dev. 2009 Dec;19(6):550-6. doi: 10.1016/j.gde.2009.10.004. Epub 2009 Oct 31. Curr Opin Genet Dev. 2009. PMID: 19880310 Free PMC article. Review.
Cited by
-
Dosage compensation in Drosophila melanogaster: epigenetic fine-tuning of chromosome-wide transcription.Nat Rev Genet. 2012 Jan 18;13(2):123-34. doi: 10.1038/nrg3124. Nat Rev Genet. 2012. PMID: 22251873 Review.
-
Genome-wide phosphoacetylation of histone H3 at Drosophila enhancers and promoters.Genome Res. 2012 Jun;22(6):1081-8. doi: 10.1101/gr.136929.111. Epub 2012 Apr 16. Genome Res. 2012. PMID: 22508764 Free PMC article.
-
Chromatin insulator factors involved in long-range DNA interactions and their role in the folding of the Drosophila genome.PLoS Genet. 2014 Aug 28;10(8):e1004544. doi: 10.1371/journal.pgen.1004544. eCollection 2014 Aug. PLoS Genet. 2014. PMID: 25165871 Free PMC article.
-
Roles of chromatin insulators in the formation of long-range contacts.Nucleus. 2015;6(2):118-22. doi: 10.1080/19491034.2015.1010962. Epub 2015 Mar 17. Nucleus. 2015. PMID: 25781057 Free PMC article. Review.
-
Targeting of Painting of fourth to roX1 and roX2 proximal sites suggests evolutionary links between dosage compensation and the regulation of the fourth chromosome in Drosophila melanogaster.G3 (Bethesda). 2013 Aug 7;3(8):1325-34. doi: 10.1534/g3.113.006866. G3 (Bethesda). 2013. PMID: 23733888 Free PMC article.
References
-
- Wang Y, Zhang W, Jin Y, Johansen J, Johansen KM. The JIL-1 tandem kinase mediates histone H3 phosphorylation and is required for maintenance of chromatin structure in Drosophila. Cell. 2001;105:433–443. - PubMed
-
- Jin Y, Wang Y, Walker DL, Dong H, Conley C, et al. JIL-1: a novel chromosomal tandem kinase implicated in transcriptional regulation in Drosophila. Mol Cell. 1999;4:129–135. - PubMed
-
- Deng H, Zhang W, Bao X, Martin JN, Girton J, et al. The JIL-1 kinase regulates the structure of Drosophila polytene chromosomes. Chromosoma. 2005;114:173–182. - PubMed
-
- Deng H, Bao X, Cai W, Blacketer MJ, Belmont AS, et al. Ectopic histone H3S10 phosphorylation causes chromatin structure remodeling in Drosophila. Development. 2008;135:699–705. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
