

The SUVR4 histone lysine methyltransferase binds ubiquitin and converts H3K9me1 to H3K9me3 on transposon chromatin in Arabidopsis
- PMID: 21423664
- PMCID: PMC3053343
- DOI: 10.1371/journal.pgen.1001325
The SUVR4 histone lysine methyltransferase binds ubiquitin and converts H3K9me1 to H3K9me3 on transposon chromatin in Arabidopsis
Abstract
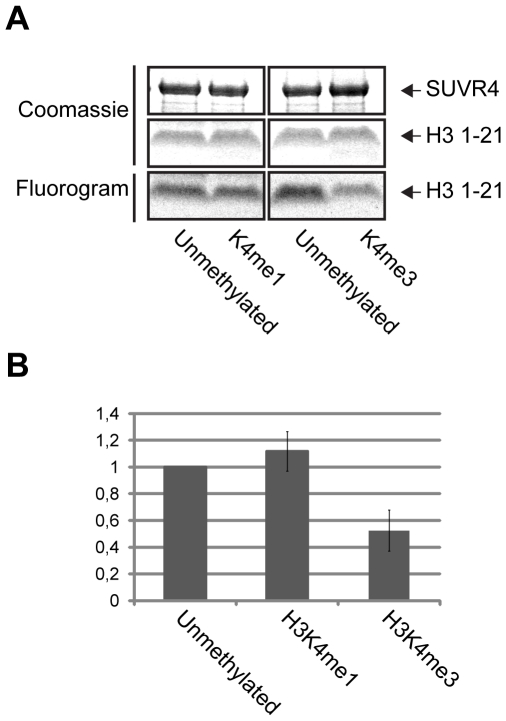
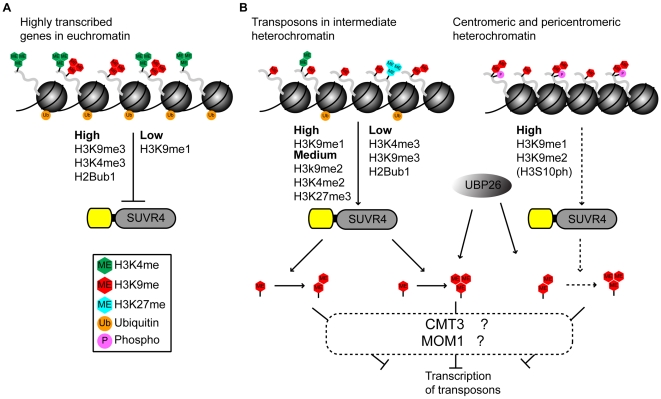
Chromatin structure and gene expression are regulated by posttranslational modifications (PTMs) on the N-terminal tails of histones. Mono-, di-, or trimethylation of lysine residues by histone lysine methyltransferases (HKMTases) can have activating or repressive functions depending on the position and context of the modified lysine. In Arabidopsis, trimethylation of lysine 9 on histone H3 (H3K9me3) is mainly associated with euchromatin and transcribed genes, although low levels of this mark are also detected at transposons and repeat sequences. Besides the evolutionarily conserved SET domain which is responsible for enzyme activity, most HKMTases also contain additional domains which enable them to respond to other PTMs or cellular signals. Here we show that the N-terminal WIYLD domain of the Arabidopsis SUVR4 HKMTase binds ubiquitin and that the SUVR4 product specificity shifts from di- to trimethylation in the presence of free ubiquitin, enabling conversion of H3K9me1 to H3K9me3 in vitro. Chromatin immunoprecipitation and immunocytological analysis showed that SUVR4 in vivo specifically converts H3K9me1 to H3K9me3 at transposons and pseudogenes and has a locus-specific repressive effect on the expression of such elements. Bisulfite sequencing indicates that this repression involves both DNA methylation-dependent and -independent mechanisms. Transcribed genes with high endogenous levels of H3K4me3, H3K9me3, and H2Bub1, but low H3K9me1, are generally unaffected by SUVR4 activity. Our results imply that SUVR4 is involved in the epigenetic defense mechanism by trimethylating H3K9 to suppress potentially harmful transposon activity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





Similar articles
-
The arabidopsis histone methyltransferase SUVR4 binds ubiquitin via a domain with a four-helix bundle structure.Biochemistry. 2014 Apr 8;53(13):2091-100. doi: 10.1021/bi401436h. Epub 2014 Mar 25. Biochemistry. 2014. PMID: 24625295 Free PMC article.
-
The Arabidopsis SUVR4 protein is a nucleolar histone methyltransferase with preference for monomethylated H3K9.Nucleic Acids Res. 2006;34(19):5461-70. doi: 10.1093/nar/gkl687. Epub 2006 Oct 4. Nucleic Acids Res. 2006. PMID: 17020925 Free PMC article.
-
SET DOMAIN GROUP25 encodes a histone methyltransferase and is involved in FLOWERING LOCUS C activation and repression of flowering.Plant Physiol. 2009 Nov;151(3):1476-85. doi: 10.1104/pp.109.143941. Epub 2009 Sep 2. Plant Physiol. 2009. PMID: 19726574 Free PMC article.
-
SET domain proteins in plant development.Biochim Biophys Acta. 2011 Aug;1809(8):407-20. doi: 10.1016/j.bbagrm.2011.05.008. Epub 2011 May 23. Biochim Biophys Acta. 2011. PMID: 21664308 Review.
-
The many faces of histone lysine methylation.Curr Opin Cell Biol. 2002 Jun;14(3):286-98. doi: 10.1016/s0955-0674(02)00335-6. Curr Opin Cell Biol. 2002. PMID: 12067650 Review.
Cited by
-
Writing and Reading Histone H3 Lysine 9 Methylation in Arabidopsis.Front Plant Sci. 2020 May 6;11:452. doi: 10.3389/fpls.2020.00452. eCollection 2020. Front Plant Sci. 2020. PMID: 32435252 Free PMC article. Review.
-
Arabidopsis histone H3 lysine 9 methyltransferases KYP/SUVH5/6 are involved in leaf development by interacting with AS1-AS2 to repress KNAT1 and KNAT2.Commun Biol. 2023 Feb 24;6(1):219. doi: 10.1038/s42003-023-04607-6. Commun Biol. 2023. PMID: 36828846 Free PMC article.
-
The ASH1-RELATED3 SET-domain protein controls cell division competence of the meristem and the quiescent center of the Arabidopsis primary root.Plant Physiol. 2014 Oct;166(2):632-43. doi: 10.1104/pp.114.244798. Epub 2014 Jul 17. Plant Physiol. 2014. PMID: 25034019 Free PMC article.
-
Inhibition of DNA methylation alters chromatin organization, nuclear positioning and activity of 45S rDNA loci in cycling cells of Q. robur.PLoS One. 2014 Aug 5;9(8):e103954. doi: 10.1371/journal.pone.0103954. eCollection 2014. PLoS One. 2014. PMID: 25093501 Free PMC article.
-
Histone methyltransferases regulating rRNA gene dose and dosage control in Arabidopsis.Genes Dev. 2012 May 1;26(9):945-57. doi: 10.1101/gad.182865.111. Genes Dev. 2012. PMID: 22549957 Free PMC article.
References
-
- Kouzarides T. Chromatin modifications and their function. Cell. 2007;128:693–705. - PubMed
-
- Latham JA, Dent SY. Cross-regulation of histone modifications. Nat Struct Mol Biol. 2007;14:1017–1024. - PubMed
-
- Yap KL, Zhou MM. Structure and function of protein modules in chromatin biology. Results Probl Cell Differ. 2006;41:1–23. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
