

Genome-wide analysis reveals the vacuolar pH-stat of Saccharomyces cerevisiae
- PMID: 21423800
- PMCID: PMC3056714
- DOI: 10.1371/journal.pone.0017619
Genome-wide analysis reveals the vacuolar pH-stat of Saccharomyces cerevisiae
Abstract
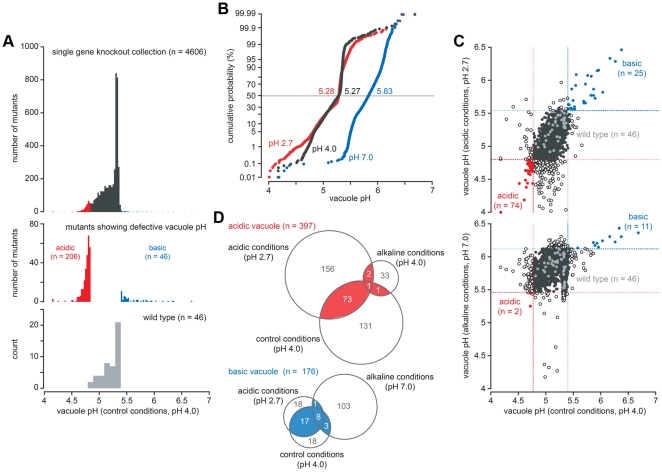
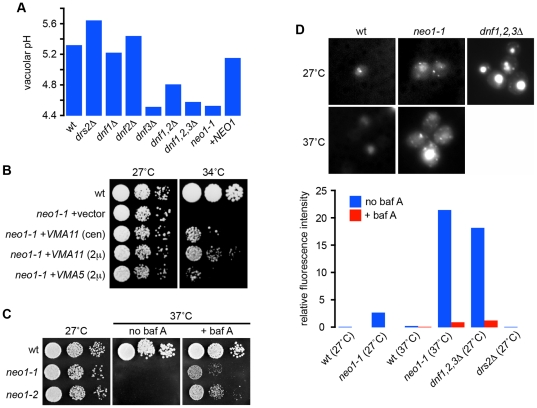
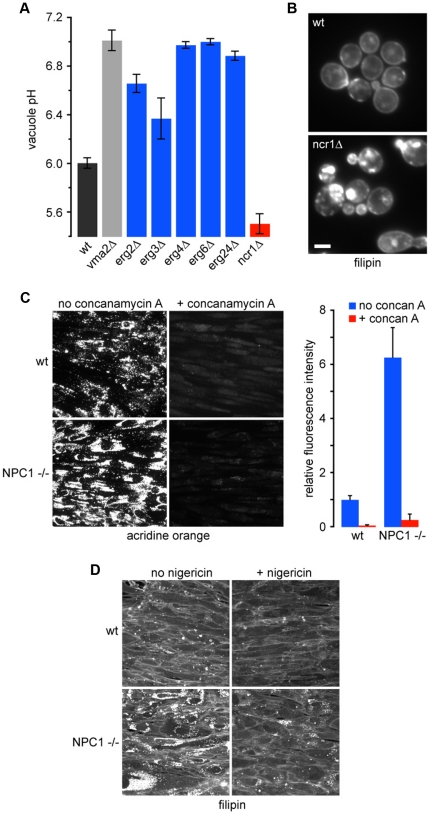
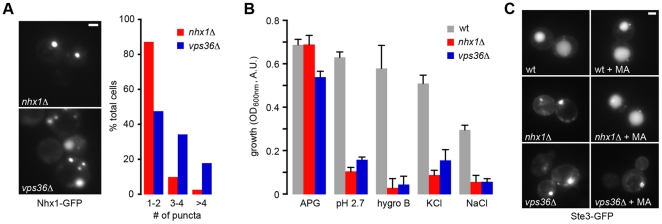
Protons, the smallest and most ubiquitous of ions, are central to physiological processes. Transmembrane proton gradients drive ATP synthesis, metabolite transport, receptor recycling and vesicle trafficking, while compartmental pH controls enzyme function. Despite this fundamental importance, the mechanisms underlying pH homeostasis are not entirely accounted for in any organelle or organism. We undertook a genome-wide survey of vacuole pH (pH(v)) in 4,606 single-gene deletion mutants of Saccharomyces cerevisiae under control, acid and alkali stress conditions to reveal the vacuolar pH-stat. Median pH(v) (5.27±0.13) was resistant to acid stress (5.28±0.14) but shifted significantly in response to alkali stress (5.83±0.13). Of 107 mutants that displayed aberrant pH(v) under more than one external pH condition, functional categories of transporters, membrane biogenesis and trafficking machinery were significantly enriched. Phospholipid flippases, encoded by the family of P4-type ATPases, emerged as pH regulators, as did the yeast ortholog of Niemann Pick Type C protein, implicated in sterol trafficking. An independent genetic screen revealed that correction of pH(v) dysregulation in a neo1(ts) mutant restored viability whereas cholesterol accumulation in human NPC1(-/-) fibroblasts diminished upon treatment with a proton ionophore. Furthermore, while it is established that lumenal pH affects trafficking, this study revealed a reciprocal link with many mutants defective in anterograde pathways being hyperacidic and retrograde pathway mutants with alkaline vacuoles. In these and other examples, pH perturbations emerge as a hitherto unrecognized phenotype that may contribute to the cellular basis of disease and offer potential therapeutic intervention through pH modulation.
Conflict of interest statement
Figures
References
-
- Kaufmann SH. Immunology's foundation: the 100-year anniversary of the Nobel Prize to Paul Ehrlich and Elie Metchnikoff. Nat Immunol. 2008;9:705–712. - PubMed
-
- de Duve C. The lysosome turns fifty. Nat Cell Biol. 2005;7:847–849. - PubMed
-
- Mellman I. The importance of being acid: the role of acidification in intracellular membrane traffic. J Exp Biol. 1992;172:39–45. - PubMed
-
- Maranda B, Brown D, Bourgoin S, Casanova JE, Vinay P, et al. Intra-endosomal pH-sensitive recruitment of the Arf-nucleotide exchange factor ARNO and Arf6 from cytoplasm to proximal tubule endosomes. J Biol Chem. 2001;276:18540–18550. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
