

Using artificial systems to explore the ecology and evolution of symbioses
- PMID: 21424911
- PMCID: PMC11114700
- DOI: 10.1007/s00018-011-0649-y
Using artificial systems to explore the ecology and evolution of symbioses
Abstract
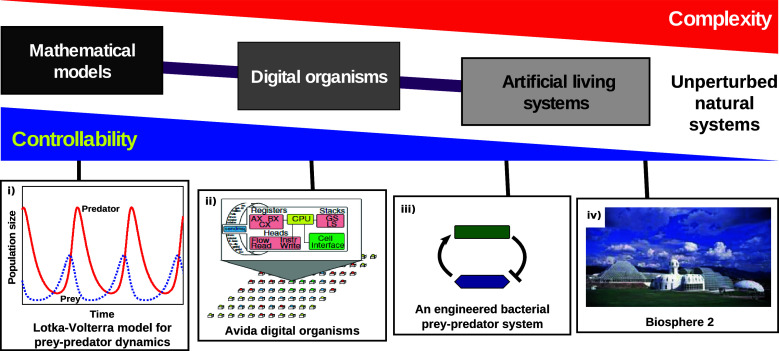
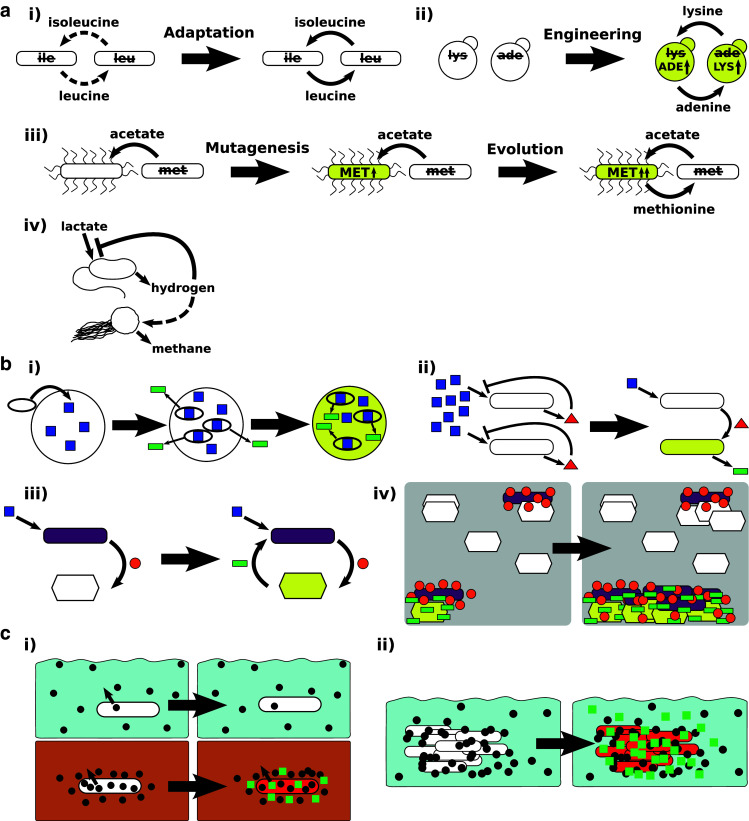
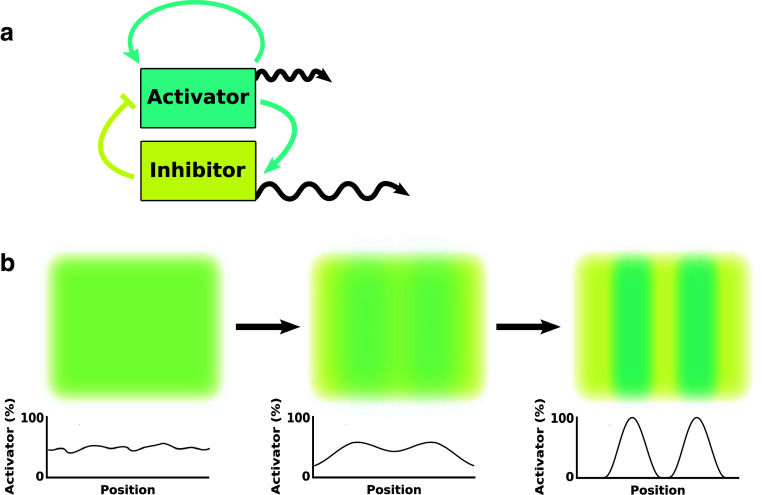
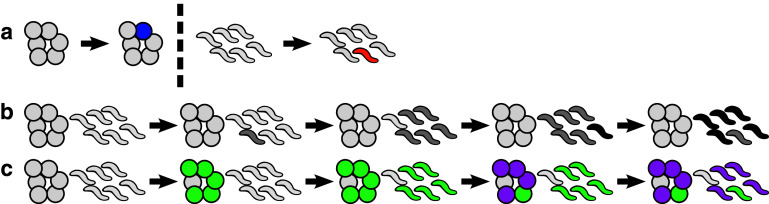
The web of life is weaved from diverse symbiotic interactions between species. Symbioses vary from antagonistic interactions such as competition and predation to beneficial interactions such as mutualism. What are the bases for the origin and persistence of symbiosis? What affects the ecology and evolution of symbioses? How do symbiotic interactions generate ecological patterns? How do symbiotic partners evolve and coevolve? Many of these questions are difficult to address in natural systems. Artificial systems, from abstract to living, have been constructed to capture essential features of natural symbioses and to address these key questions. With reduced complexity and increased controllability, artificial systems can serve as useful models for natural systems. We review how artificial systems have contributed to our understanding of symbioses.
Figures
Similar articles
-
Current issues in the evolutionary ecology of ant-plant symbioses.New Phytol. 2014 May;202(3):749-764. doi: 10.1111/nph.12690. Epub 2014 Jan 21. New Phytol. 2014. PMID: 24444030 Review.
-
Simulation of evolution implemented in the mutualistic symbioses towards enhancing their ecological efficiency, functional integrity and genotypic specificity.Theor Popul Biol. 2010 Dec;78(4):259-69. doi: 10.1016/j.tpb.2010.08.005. Epub 2010 Sep 9. Theor Popul Biol. 2010. PMID: 20832414
-
The role of exploitation in the establishment of mutualistic microbial symbioses.FEMS Microbiol Lett. 2019 Jun 1;366(12):fnz148. doi: 10.1093/femsle/fnz148. FEMS Microbiol Lett. 2019. PMID: 31271421 Free PMC article.
-
The roles of amensalistic and commensalistic interactions in large ecological network stability.Sci Rep. 2016 Jul 13;6:29929. doi: 10.1038/srep29929. Sci Rep. 2016. PMID: 27406267 Free PMC article.
-
A mechanistic framework for complex microbe-host symbioses.Trends Microbiol. 2025 Jan;33(1):96-111. doi: 10.1016/j.tim.2024.08.002. Epub 2024 Sep 5. Trends Microbiol. 2025. PMID: 39242229 Review.
Cited by
-
Synthetic Symbiosis under Environmental Disturbances.mSystems. 2020 Jun 16;5(3):e00187-20. doi: 10.1128/mSystems.00187-20. mSystems. 2020. PMID: 32546669 Free PMC article.
-
Enhanced metabolic entanglement emerges during the evolution of an interkingdom microbial community.Nat Commun. 2024 Aug 22;15(1):7238. doi: 10.1038/s41467-024-51702-1. Nat Commun. 2024. PMID: 39174531 Free PMC article.
-
Niche differentiation among comammox (Nitrospira inopinata) and other metabolically distinct nitrifiers.Front Microbiol. 2022 Sep 14;13:956860. doi: 10.3389/fmicb.2022.956860. eCollection 2022. Front Microbiol. 2022. PMID: 36187961 Free PMC article.
-
Spatial structure favors microbial coexistence except when slower mediator diffusion weakens interactions.Elife. 2023 Jun 23;12:e82504. doi: 10.7554/eLife.82504. Elife. 2023. PMID: 37350317 Free PMC article.
-
Spatial dynamics of synthetic microbial mutualists and their parasites.PLoS Comput Biol. 2017 Aug 21;13(8):e1005689. doi: 10.1371/journal.pcbi.1005689. eCollection 2017 Aug. PLoS Comput Biol. 2017. PMID: 28827802 Free PMC article.
References
-
- Wilkinson D. At cross purposes. Nature. 2001;412:485. - PubMed
-
- Boucher D (1985) In: Boucher (D) (ed) The biology of mutualism: ecology and evolution. Oxford University Press, New York
-
- Neutel A, Heesterbeek J, van de Koppel J, Hoenderboom G, Vos A, Kaldeway C, Berendse F, de Ruiter P. Reconciling complexity with stability in naturally assembling food webs. Nature. 2007;449:599–602. - PubMed
-
- Tilman D, Kareiva P. Spatial ecology. USA: Princeton University Press; 1997.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
