

COP1-mediated degradation of BBX22/LZF1 optimizes seedling development in Arabidopsis
- PMID: 21427283
- PMCID: PMC3091042
- DOI: 10.1104/pp.111.175042
COP1-mediated degradation of BBX22/LZF1 optimizes seedling development in Arabidopsis
Abstract
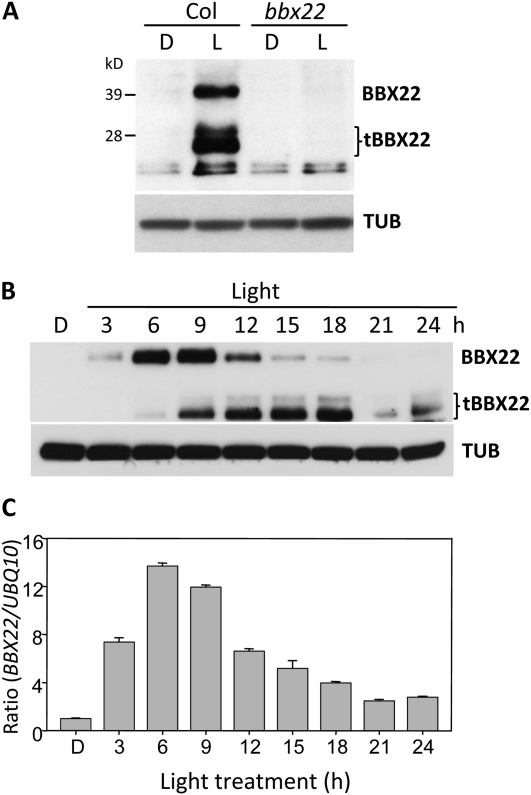
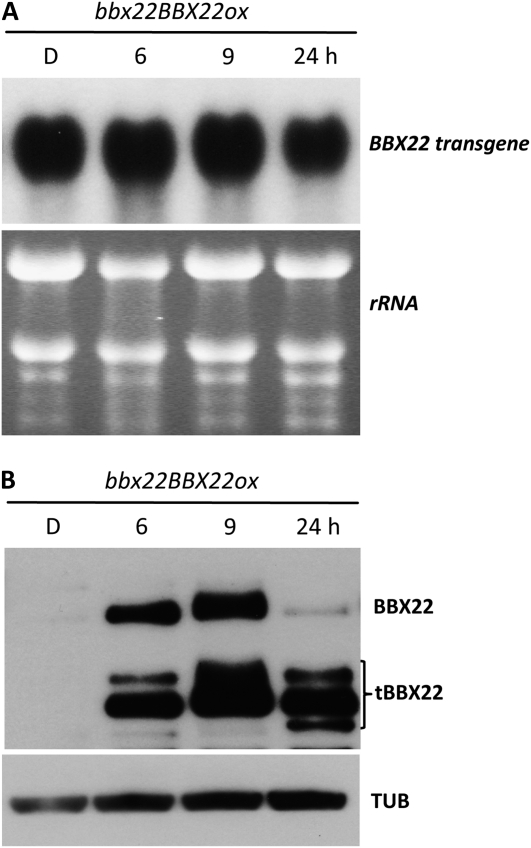
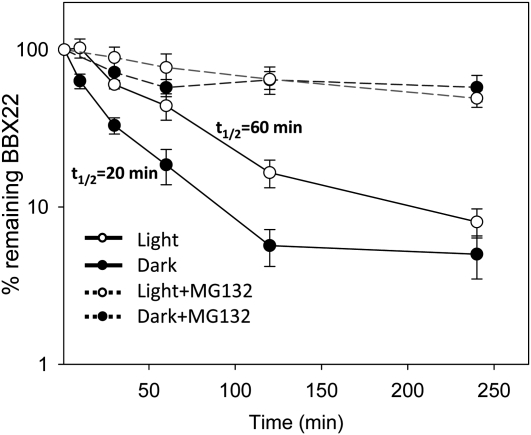
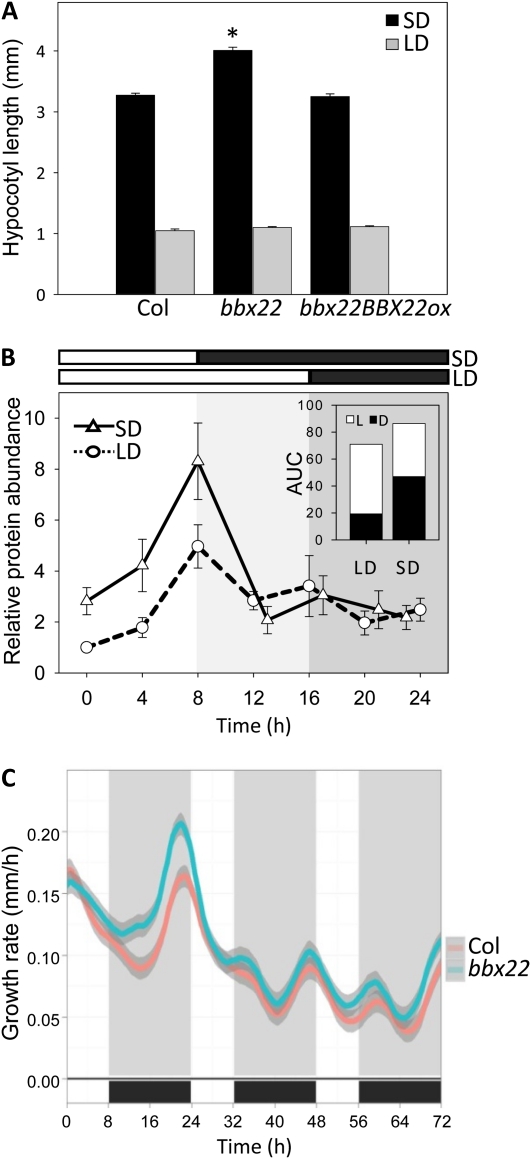
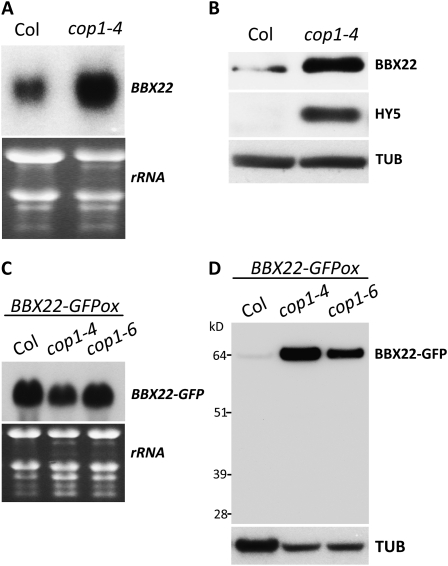
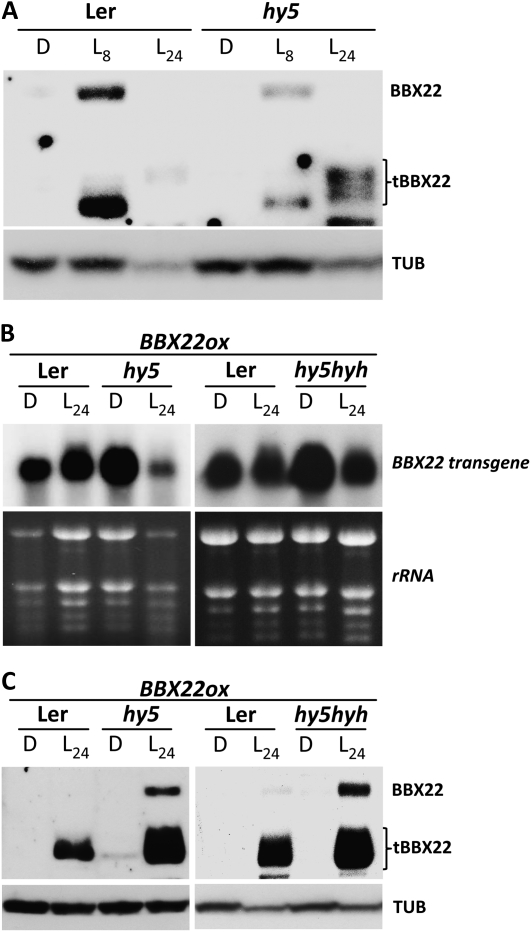
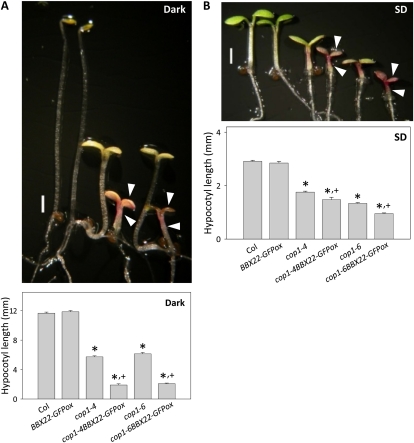
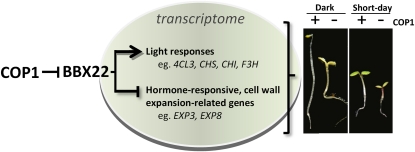
Light regulates multiple aspects of growth and development in plants. Transcriptomic changes govern the expression of signaling molecules with the perception of light. Also, the 26S proteasome regulates the accumulation of positive and negative regulators for optimal growth of Arabidopsis (Arabidopsis thaliana) in the dark, light, or light/dark cycles. BBX22, whose induction is both light regulated and HY5 dependent, is a positive regulator of deetiolation in Arabidopsis. We found that during skotomorphogenesis, the expression of BBX22 needs to be tightly regulated at both transcriptional and posttranslational levels. During photomorphogenesis, the expression of BBX22 transiently accumulates to execute its roles as a positive regulator. BBX22 protein accumulates to a higher level under short-day conditions and functions to inhibit hypocotyl elongation. The proteasome-dependent degradation of BBX22 protein is tightly controlled even in plants overexpressing BBX22. An analysis of BBX22 degradation kinetics shows that the protein has a short half-life under both dark and light conditions. COP1 mediates the degradation of BBX22 in the dark. Although dispensable in the dark, HY5 contributes to the degradation of BBX22 in the light. The constitutive photomorphogenic development of the cop1 mutant is enhanced in cop1BBX22ox plants, which show a short hypocotyl, high anthocyanin accumulation, and expression of light-responsive genes. Exaggerated light responsiveness is also observed in cop1BBX22ox seedlings grown under short-day conditions. Therefore, the proper accumulation of BBX22 is crucial for plants to maintain optimal growth when grown in the dark as well as to respond to seasonal changes in daylength.
Figures
References
-
- Agresti A. (1992) A survey of exact inference for contingency tables. Stat Sci 7: 131–153
-
- Alabadí D, Gallego-Bartolomé J, Orlando L, García-Cárcel L, Rubio V, Martínez C, Frigerio M, Iglesias-Pedraz JM, Espinosa A, Deng XW, et al. (2008) Gibberellins modulate light signaling pathways to prevent Arabidopsis seedling de-etiolation in darkness. Plant J 53: 324–335 - PubMed
-
- Al-Sady B, Ni W, Kircher S, Schäfer E, Quail PH. (2006) Photoactivated phytochrome induces rapid PIF3 phosphorylation prior to proteasome-mediated degradation. Mol Cell 23: 439–446 - PubMed
-
- Ang LH, Chattopadhyay S, Wei N, Oyama T, Okada K, Batschauer A, Deng XW. (1998) Molecular interaction between COP1 and HY5 defines a regulatory switch for light control of Arabidopsis development. Mol Cell 1: 213–222 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
