

Thymic T-cell development in allogeneic stem cell transplantation
- PMID: 21427289
- PMCID: PMC3128475
- DOI: 10.1182/blood-2011-02-334623
Thymic T-cell development in allogeneic stem cell transplantation
Abstract
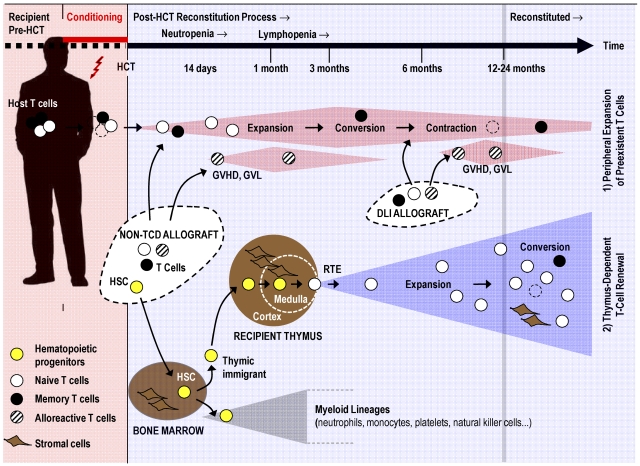
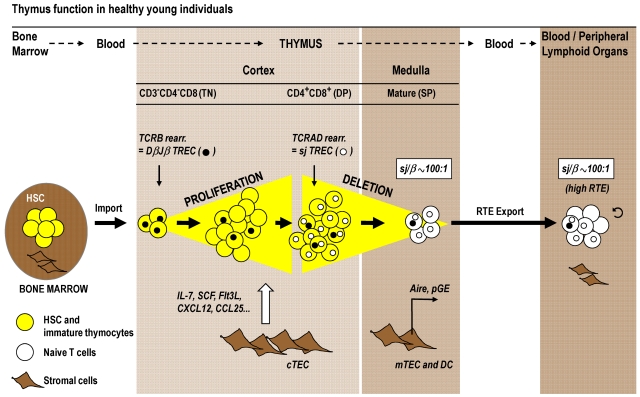
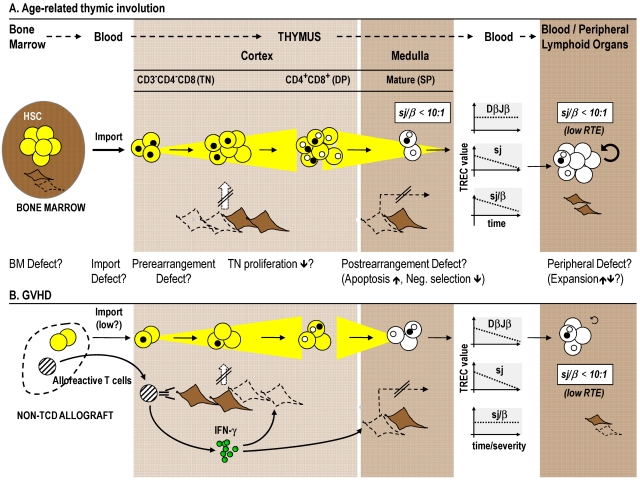
Cytoreductive conditioning regimens used in the context of allogeneic hematopoietic cell transplantation (HCT) elicit deficits in innate and adaptive immunity, which predispose patients to infections. As such, transplantation outcomes depend vitally on the successful reconstruction of immune competence. Restoration of a normal peripheral T-cell pool after HCT is a slow process that requires the de novo production of naive T cells in a functionally competent thymus. However, there are several challenges to this regenerative process. Most notably, advanced age, the cytotoxic pretransplantation conditioning, and posttransplantation alloreactivity are risk factors for T-cell immune deficiency as they independently interfere with normal thymus function. Here, we discuss preclinical allogeneic HCT models and clinical observations that have contributed to a better understanding of the transplant-related thymic dysfunction. The identification of the cellular and molecular mechanisms that control regular thymopoiesis but are altered in HCT patients is expected to provide the basis for new therapies that improve the regeneration of the adaptive immune system, especially with functionally competent, naive T cells.
Figures
References
-
- Gratwohl A, Baldomero H. Trends of hematopoietic stem cell transplantation in the third millennium. Curr Opin Hematol. 2009;16(6):420–426. - PubMed
-
- Ballen KK, King RJ, Chitphakdithai P, et al. The national marrow donor program: 20 years of unrelated donor hematopoietic cell transplantation. Biol Blood Marrow Transplant. 2008;14(9 suppl):2–7. - PubMed
-
- Pasquini MC, Wang Z. Current use and outcome of hematopoietic stem cell transplantation: II. CIBMTR Summary Slides 2009. CIBMTR Newsletter. 2009;15(2):7–11.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
