

Effects and mechanisms of wakefulness on local cortical networks
- PMID: 21435553
- PMCID: PMC3069934
- DOI: 10.1016/j.neuron.2011.02.040
Effects and mechanisms of wakefulness on local cortical networks
Abstract
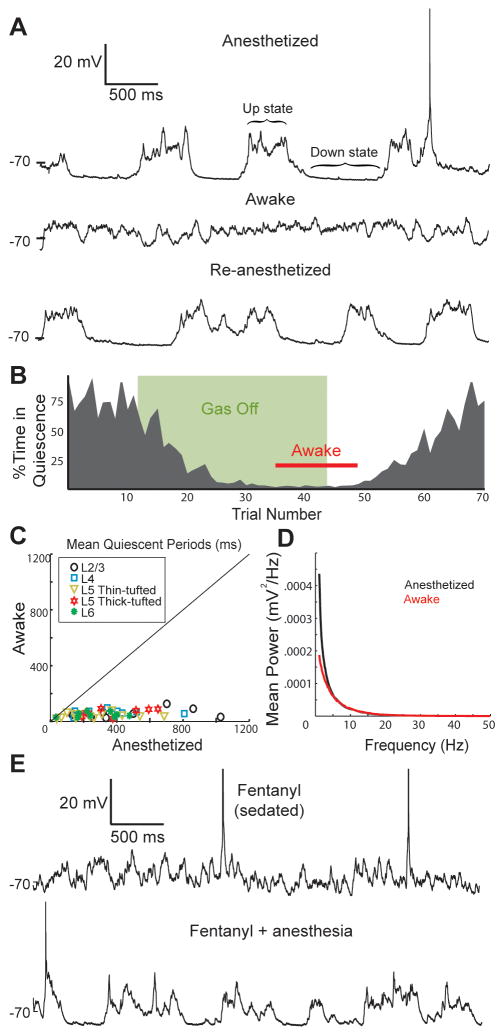
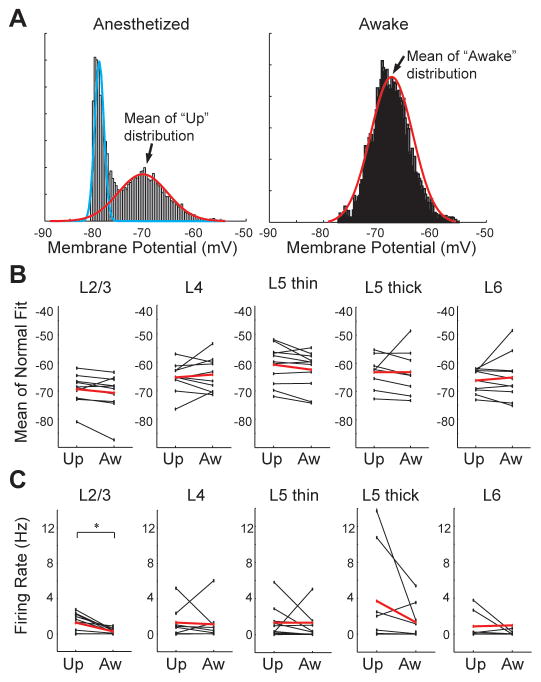
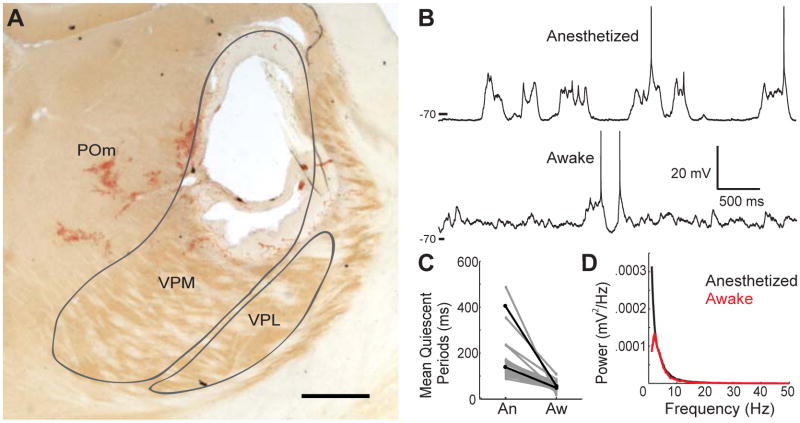
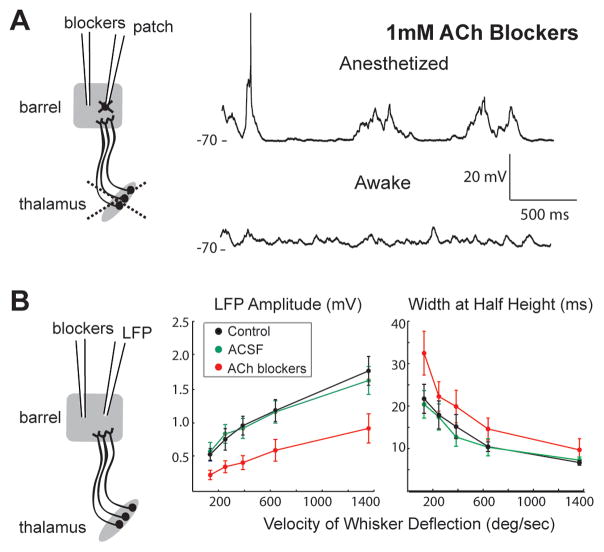
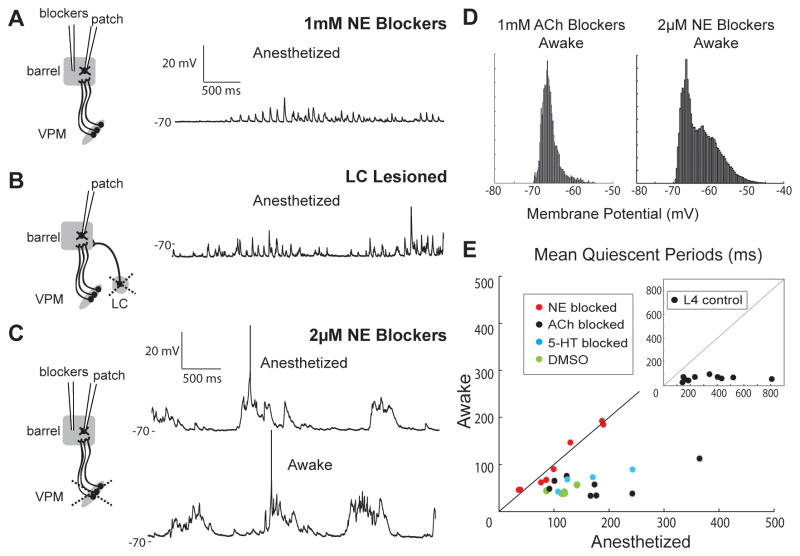
Mammalian brains generate internal activity independent of environmental stimuli. Internally generated states may bring about distinct cortical processing modes. To investigate how brain state impacts cortical circuitry, we recorded intracellularly from the same neurons, under anesthesia and subsequent wakefulness, in rat barrel cortex. In every cell examined throughout layers 2-6, wakefulness produced a temporal pattern of synaptic inputs differing markedly from those under anesthesia. Recurring periods of synaptic quiescence, prominent under anesthesia, were abolished by wakefulness, which produced instead a persistently depolarized state. This switch in dynamics was unaffected by elimination of afferent synaptic input from thalamus, suggesting that arousal alters cortical dynamics by neuromodulators acting directly on cortex. Indeed, blockade of noradrenergic, but not cholinergic, pathways induced synaptic quiescence during wakefulness. We conclude that global brain states can switch local recurrent networks into different regimes via direct neuromodulation.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Berridge CW, Page ME, Valentino RJ, Foote SL. Effects of locus coeruleus inactivation on electroencephalographic activity in neocortex and hippocampus. Neuroscience. 1993;55:381–393. - PubMed
-
- Blake H, Gerard RW. Brain potential during sleep. American Journal of Physiology. 1937;119:692–702.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
