

The maturation factors HoxR and HoxT contribute to oxygen tolerance of membrane-bound [NiFe] hydrogenase in Ralstonia eutropha H16
- PMID: 21441514
- PMCID: PMC3133145
- DOI: 10.1128/JB.01427-10
The maturation factors HoxR and HoxT contribute to oxygen tolerance of membrane-bound [NiFe] hydrogenase in Ralstonia eutropha H16
Abstract
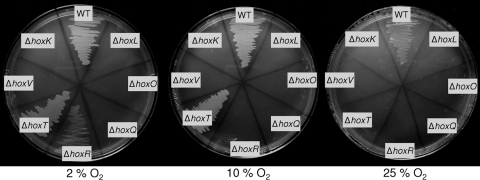
The membrane-bound [NiFe] hydrogenase (MBH) of Ralstonia eutropha H16 undergoes a complex maturation process comprising cofactor assembly and incorporation, subunit oligomerization, and finally twin-arginine-dependent membrane translocation. Due to its outstanding O(2) and CO tolerance, the MBH is of biotechnological interest and serves as a molecular model for a robust hydrogen catalyst. Adaptation of the enzyme to oxygen exposure has to take into account not only the catalytic reaction but also biosynthesis of the intricate redox cofactors. Here, we report on the role of the MBH-specific accessory proteins HoxR and HoxT, which are key components in MBH maturation at ambient O(2) levels. MBH-driven growth on H(2) is inhibited or retarded at high O(2) partial pressure (pO(2)) in mutants inactivated in the hoxR and hoxT genes. The ratio of mature and nonmature forms of the MBH small subunit is shifted toward the precursor form in extracts derived from the mutant cells grown at high pO(2). Lack of hoxR and hoxT can phenotypically be restored by providing O(2)-limited growth conditions. Analysis of copurified maturation intermediates leads to the conclusion that the HoxR protein is a constituent of a large transient protein complex, whereas the HoxT protein appears to function at a final stage of MBH maturation. UV-visible spectroscopy of heterodimeric MBH purified from hoxR mutant cells points to alterations of the Fe-S cluster composition. Thus, HoxR may play a role in establishing a specific Fe-S cluster profile, whereas the HoxT protein seems to be beneficial for cofactor stability under aerobic conditions.
Figures







Similar articles
-
Rubredoxin-related maturation factor guarantees metal cofactor integrity during aerobic biosynthesis of membrane-bound [NiFe] hydrogenase.J Biol Chem. 2014 Mar 14;289(11):7982-93. doi: 10.1074/jbc.M113.544668. Epub 2014 Jan 21. J Biol Chem. 2014. PMID: 24448806 Free PMC article.
-
Chaperones specific for the membrane-bound [NiFe]-hydrogenase interact with the Tat signal peptide of the small subunit precursor in Ralstonia eutropha H16.Mol Microbiol. 2007 Oct;66(2):453-67. doi: 10.1111/j.1365-2958.2007.05933.x. Epub 2007 Sep 10. Mol Microbiol. 2007. PMID: 17850259
-
[NiFe] and [FeS] cofactors in the membrane-bound hydrogenase of Ralstonia eutropha investigated by X-ray absorption spectroscopy: insights into O(2)-tolerant H(2) cleavage.Biochemistry. 2011 Jul 5;50(26):5858-69. doi: 10.1021/bi200367u. Epub 2011 Jun 10. Biochemistry. 2011. PMID: 21618994
-
[NiFe]-hydrogenases of Ralstonia eutropha H16: modular enzymes for oxygen-tolerant biological hydrogen oxidation.J Mol Microbiol Biotechnol. 2005;10(2-4):181-96. doi: 10.1159/000091564. J Mol Microbiol Biotechnol. 2005. PMID: 16645314 Review.
-
H2 conversion in the presence of O2 as performed by the membrane-bound [NiFe]-hydrogenase of Ralstonia eutropha.Chemphyschem. 2010 Apr 26;11(6):1107-19. doi: 10.1002/cphc.200901002. Chemphyschem. 2010. PMID: 20186906 Review.
Cited by
-
One pot synthesis of two cobalt(iii) Schiff base complexes with chelating pyridyltetrazolate and exploration of their bio-relevant catalytic activities.RSC Adv. 2018 Aug 7;8(49):28216-28237. doi: 10.1039/c8ra03035a. eCollection 2018 Aug 2. RSC Adv. 2018. PMID: 35542722 Free PMC article.
-
Distribution of Hydrogenases in Cyanobacteria: A Phylum-Wide Genomic Survey.Front Genet. 2016 Dec 27;7:223. doi: 10.3389/fgene.2016.00223. eCollection 2016. Front Genet. 2016. PMID: 28083017 Free PMC article.
-
Rubredoxin-related maturation factor guarantees metal cofactor integrity during aerobic biosynthesis of membrane-bound [NiFe] hydrogenase.J Biol Chem. 2014 Mar 14;289(11):7982-93. doi: 10.1074/jbc.M113.544668. Epub 2014 Jan 21. J Biol Chem. 2014. PMID: 24448806 Free PMC article.
-
Inactivation of the uptake hydrogenase in the purple non-sulfur photosynthetic bacterium Rubrivivax gelatinosus CBS enables a biological water-gas shift platform for H2 production.J Ind Microbiol Biotechnol. 2019 Jul;46(7):993-1002. doi: 10.1007/s10295-019-02173-7. Epub 2019 Apr 9. J Ind Microbiol Biotechnol. 2019. PMID: 30968274
-
Dual role of HupF in the biosynthesis of [NiFe] hydrogenase in Rhizobium leguminosarum.BMC Microbiol. 2012 Nov 8;12:256. doi: 10.1186/1471-2180-12-256. BMC Microbiol. 2012. PMID: 23136881 Free PMC article.
References
-
- Alami M., et al. 2003. Differential interactions between a twin-arginine signal peptide and its translocase in Escherichia coli. Mol. Cell 12:937–946 - PubMed
-
- Ayhan M., et al. 1996. The rubredoxin from Clostridium pasteurianum: mutation of the conserved glycine residues 10 and 43 to alanine and valine. Inorg. Chem. 35:5902–5911
-
- Bachmann J., Bauer B., Zwicker K., Ludwig B., Anderka O. 2006. The Rieske protein from Paracoccus denitrificans is inserted into the cytoplasmic membrane by the twin-arginine translocase. FEBS J. 273:4817–4830 - PubMed
-
- Baginsky C., Palacios J. M., Imperial J., Ruiz-Argueso T., Brito B. 2004. Molecular and functional characterization of the Azorhizobium caulinodans ORS571 hydrogenase gene cluster. FEMS Microbiol. Lett. 237:399–405 - PubMed
-
- Bendtsen J. D., Nielsen H., von Heijne G., Brunak S. 2004. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 340:783–795 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
