

ABC ATPase signature helices in Rad50 link nucleotide state to Mre11 interface for DNA repair
- PMID: 21441914
- PMCID: PMC3118400
- DOI: 10.1038/nsmb.2038
ABC ATPase signature helices in Rad50 link nucleotide state to Mre11 interface for DNA repair
Erratum in
- Nat Struct Mol Biol. 2011 Sep;18(9):1084
- Nat Struct Mol Biol. 2012 Mar;19(3):364
Abstract
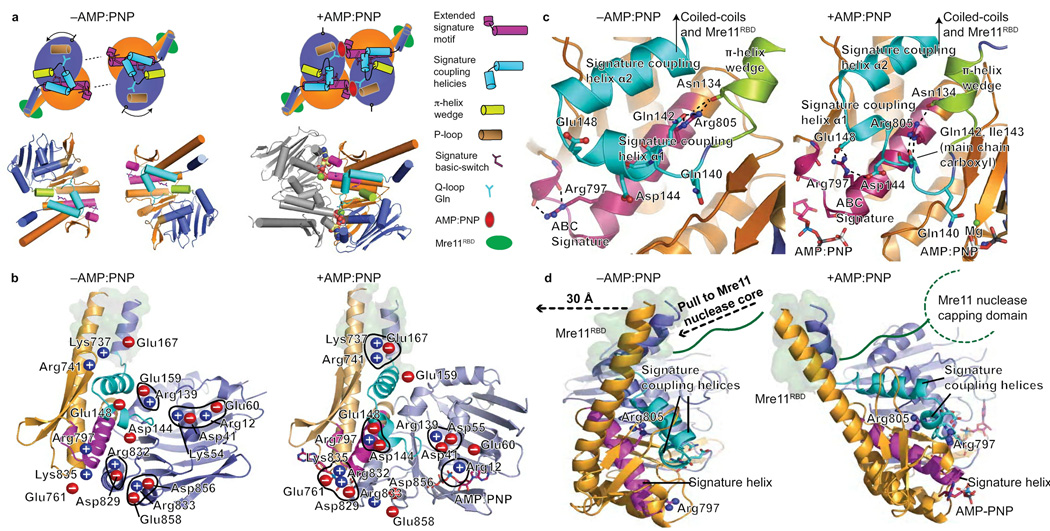
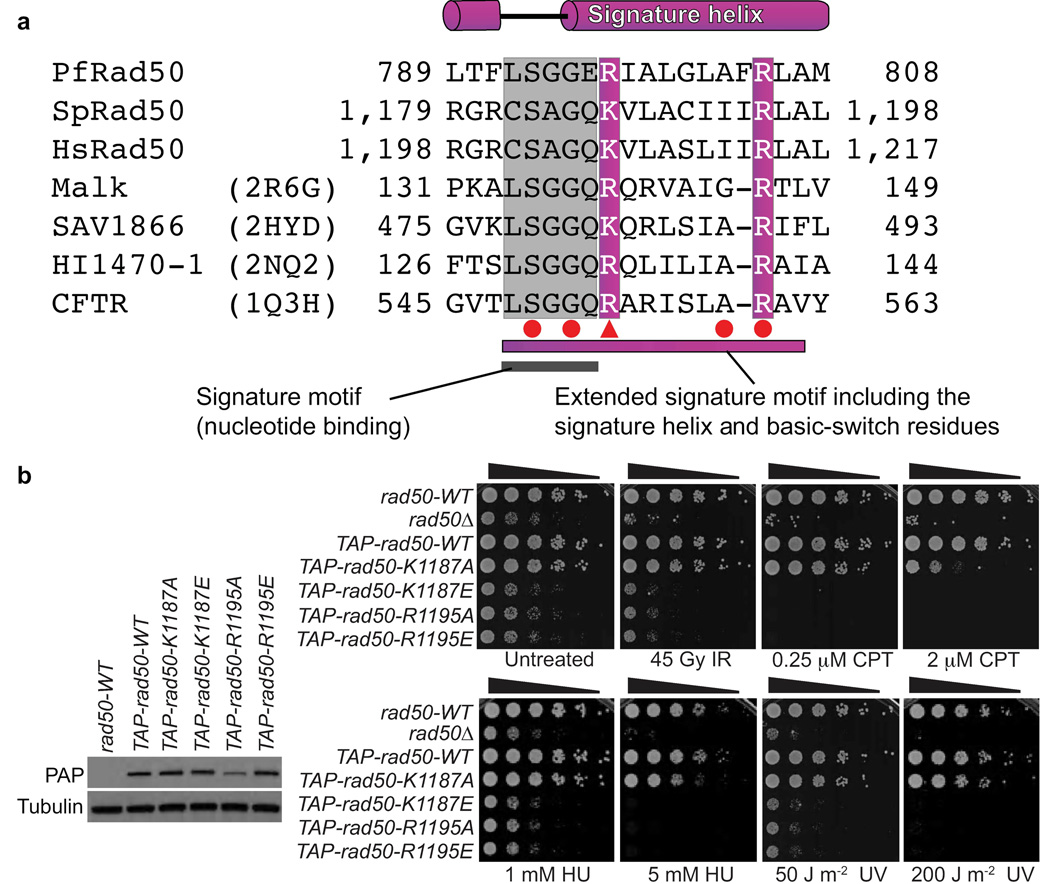
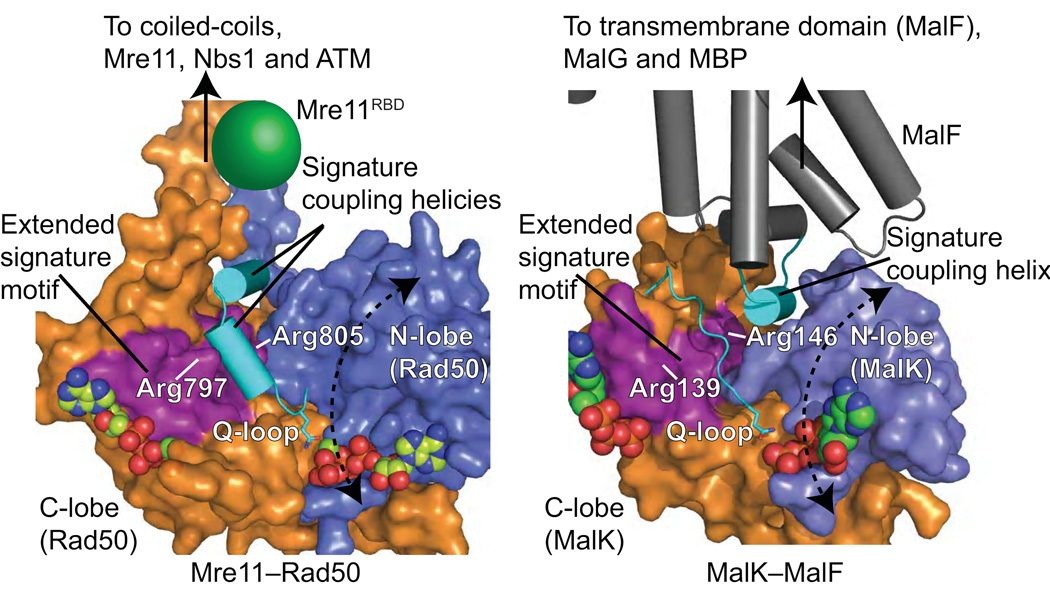
The Rad50 ABC-ATPase complex with Mre11 nuclease is essential for dsDNA break repair, telomere maintenance and ataxia telangiectasia-mutated kinase checkpoint signaling. How Rad50 affects Mre11 functions and how ABC-ATPases communicate nucleotide binding and ligand states across long distances and among protein partners are questions that have remained obscure. Here, structures of Mre11-Rad50 complexes define the Mre11 2-helix Rad50 binding domain (RBD) that forms a four-helix interface with Rad50 coiled coils adjoining the ATPase core. Newly identified effector and basic-switch helix motifs extend the ABC-ATPase signature motif to link ATP-driven Rad50 movements to coiled coils binding Mre11, implying an ~30-Å pull on the linker to the nuclease domain. Both RBD and basic-switch mutations cause clastogen sensitivity. Our new results characterize flexible ATP-dependent Mre11 regulation, defects in cancer-linked RBD mutations, conserved superfamily basic switches and motifs effecting ATP-driven conformational change, and they provide a unified comprehension of ABC-ATPase activities.
Figures
References
-
- Trujillo KM, Sung P. DNA structure-specific nuclease activities in the Saccharomyces cerevisiae Rad50-Mre11 complex. J. Biol. Chem. 2001;276:35458–35464. - PubMed
-
- Williams RS, Williams JS, Tainer JA. Mre11-Rad50-Nbs1 is a keystone complex connecting DNA repair machinery, double-strand break signaling, and the chromatin template. Biochem. Cell Biol. 2007;85:509–520. - PubMed
-
- Carney JP, et al. The hMre11/hRad50 protein complex and Nijmegen breakage syndrome: linkage of double-strand break repair to the cellular DNA damage response. Cell. 1998;93:477–486. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
