

Development and organization of the lamprey telencephalon with special reference to the GABAergic system
- PMID: 21442003
- PMCID: PMC3062466
- DOI: 10.3389/fnana.2011.00020
Development and organization of the lamprey telencephalon with special reference to the GABAergic system
Abstract
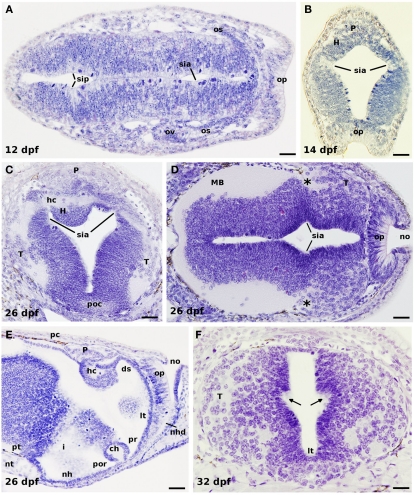
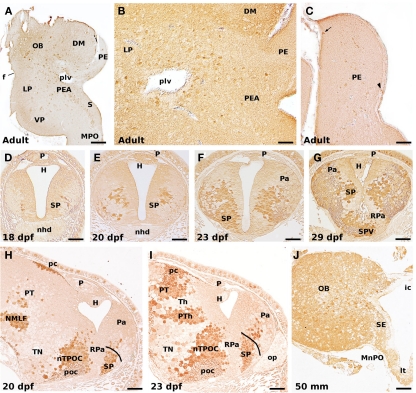
Lampreys, together with hagfishes, represent the sister group of gnathostome vertebrates. There is an increasing interest for comparing the forebrain organization observed in lampreys and gnathostomes to shed light on vertebrate brain evolution. Within the prosencephalon, there is now a general agreement on the major subdivisions of the lamprey diencephalon; however, the organization of the telencephalon, and particularly its pallial subdivisions, is still a matter of controversy. In this study, recent progress on the development and organization of the lamprey telencephalon is reviewed, with particular emphasis on the GABA immunoreactive cell populations trying to understand their putative origin. First, we describe some early general cytoarchitectonic events by searching the classical literature as well as our collection of embryonic and prolarval series of hematoxylin-stained sections. Then, we comment on the cell proliferation activity throughout the larval period, followed by a detailed description of the early events on the development of the telencephalic GABAergic system. In this context, lampreys apparently do not possess the same molecularly distinct subdivisions of the gnathostome basal telencephalon because of the absence of a Nkx2.1-expressing domain in the developing subpallium; a fact that has been related to the absence of a medial ganglionic eminence as well as of its derived nucleus in gnathostomes, the pallidum. Therefore, these data raise interesting questions such as whether or not a different mechanism to specify telencephalic GABAergic neurons exists in lampreys or what are their migration pathways. Finally, we summarize the organization of the adult lamprey telencephalon by analyzing the main proposed conceptions, including the available data on the expression pattern of some developmental regulatory genes which are of importance for building its adult shape.
Keywords: agnathans; cell proliferation; cyclostomes; evolution; olfactory bulbs; pallium; preoptic region; subpallium.
Figures




Similar articles
-
Involvement of Hedgehog and FGF signalling in the lamprey telencephalon: evolution of regionalization and dorsoventral patterning of the vertebrate forebrain.Development. 2011 Mar;138(6):1217-26. doi: 10.1242/dev.059360. Development. 2011. PMID: 21343370
-
Ontogeny of gamma-aminobutyric acid-immunoreactive neuronal populations in the forebrain and midbrain of the sea lamprey.J Comp Neurol. 2002 May 13;446(4):360-76. doi: 10.1002/cne.10209. J Comp Neurol. 2002. PMID: 11954035
-
Reconstructing the ancestral vertebrate brain.Dev Growth Differ. 2017 May;59(4):163-174. doi: 10.1111/dgd.12347. Epub 2017 Apr 26. Dev Growth Differ. 2017. PMID: 28447337 Review.
-
Telencephalic connections in the Pacific hagfish (Eptatretus stouti), with special reference to the thalamopallial system.J Comp Neurol. 1998 Jun 1;395(2):245-60. doi: 10.1002/(sici)1096-9861(19980601)395:2<245::aid-cne8>3.0.co;2-2. J Comp Neurol. 1998. PMID: 9603376
-
Forebrain Architecture and Development in Cyclostomes, with Reference to the Early Morphology and Evolution of the Vertebrate Head.Brain Behav Evol. 2022;96(4-6):305-317. doi: 10.1159/000519026. Epub 2021 Sep 17. Brain Behav Evol. 2022. PMID: 34537767 Review.
Cited by
-
Evidence from cyclostomes for complex regionalization of the ancestral vertebrate brain.Nature. 2016 Mar 3;531(7592):97-100. doi: 10.1038/nature16518. Epub 2016 Feb 15. Nature. 2016. PMID: 26878236
-
GABAergic modulation of olfactomotor transmission in lampreys.PLoS Biol. 2018 Oct 4;16(10):e2005512. doi: 10.1371/journal.pbio.2005512. eCollection 2018 Oct. PLoS Biol. 2018. PMID: 30286079 Free PMC article.
-
Integrative neuro-endocrine pathways in the control of reproduction in lamprey: a brief review.Front Endocrinol (Lausanne). 2013 Oct 18;4:151. doi: 10.3389/fendo.2013.00151. Front Endocrinol (Lausanne). 2013. PMID: 24151489 Free PMC article. Review.
-
A lamprey neural cell type atlas illuminates the origins of the vertebrate brain.Nat Ecol Evol. 2023 Oct;7(10):1714-1728. doi: 10.1038/s41559-023-02170-1. Epub 2023 Sep 14. Nat Ecol Evol. 2023. PMID: 37710042 Free PMC article.
-
Interneuron Origins in the Embryonic Porcine Medial Ganglionic Eminence.J Neurosci. 2021 Apr 7;41(14):3105-3119. doi: 10.1523/JNEUROSCI.2738-20.2021. Epub 2021 Feb 26. J Neurosci. 2021. PMID: 33637558 Free PMC article.
References
-
- Ahlborn F. (1883). Untersuchungen über das Gehirn der Petromyzonten. Z. Wissensch. Zool. XXXIX, 191–294
-
- álvarez-Otero R., Megías M., Pombal M. A. (2002). “Postembryonic cell proliferation in the sea lamprey central nervous system,” in FENS Forum, Paris (Abstr. A 137.1, 171).
-
- Bergquist H. (1952). Studies on the cerebral tube in vertebrates. The neuromeres. Acta Zool. 33, 117–18710.1111/j.1463-6395.1952.tb00362.x - DOI
-
- Brox A., Puelles L., Ferreiro B., Medina L. (2003). Expression of the genes GAD67 and Distal-less-4 in the forebrain of Xenopus laevis confirms a common pattern in tetrapods. J. Comp. Neurol. 461, 370–393 - PubMed
LinkOut - more resources
Full Text Sources