

Physiology of the gonadotrophin-releasing hormone (GnRH) neurone: studies from embryonic GnRH neurones
- PMID: 21443528
- PMCID: PMC3101116
- DOI: 10.1111/j.1365-2826.2011.02130.x
Physiology of the gonadotrophin-releasing hormone (GnRH) neurone: studies from embryonic GnRH neurones
Abstract
Gonadotrophin-releasing hormone (GnRH)-secreting neurones are the final output of the central nervous system driving fertility in all mammals. Although it has been known for decades that the efficiency of communication between the hypothalamus and the pituitary depends on the pulsatile profile of GnRH secretion, how GnRH neuronal activity is patterned to generate pulses at the median eminence is unknown. To date, the scattered distribution of the GnRH cell bodies remains the main limitation to assessing the cellular events that could lead to pulsatile GnRH secretion. Taking advantage of the unique developmental feature of GnRH neurones, the nasal explant model allows primary GnRH neurones to be maintained within a micro-network where pulsatile secretion is preserved and where individual cellular activity can be monitored simultaneously across the cell population. This review summarises the data obtained from work using this in vitro model, and brings some insights into GnRH cellular physiology.
© 2011 The Author. Journal of Neuroendocrinology © 2011 Blackwell Publishing Ltd.
Figures
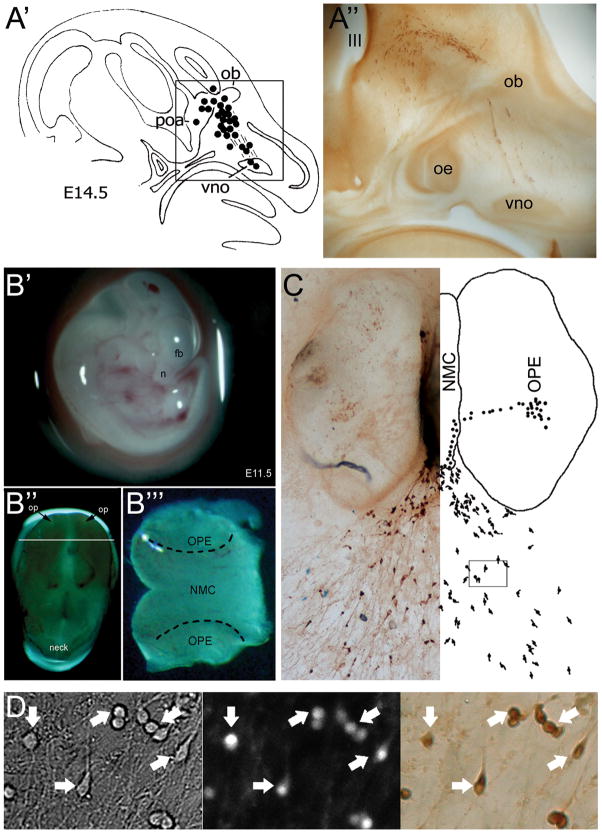



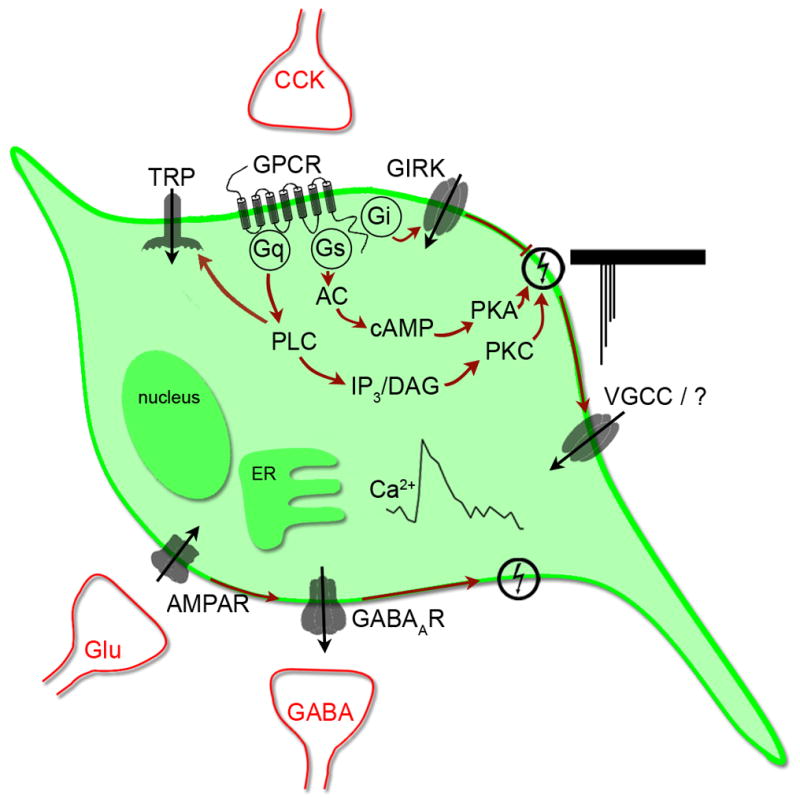
References
-
- King JC, Tobet SA, Snavely FL, Arimura AA. LHRH immunopositive cells and their projections to the median eminence and organum vasculosum of the lamina terminalis. J Comp Neurol. 1982;209(3):287–300. - PubMed
-
- Hoffman GE, Phelps CJ, Khachaturian H, Sladek JR., Jr . Neuroendocrine Projections to the Median Eminence. In: Ganten D, Pfaff D, editors. Morphology of Hypothalamus and Its Connections. Berlin - Heidelberg - New York - London -Paris - Tokyo: Academic Press; 1986. pp. 161–96.
-
- McCann SM, Ramirez VD. The Neuroendocrine Regulation of Hypophyseal Luteinizing Hormone Secretion. Recent Prog Horm Res. 1964:20131–81. - PubMed
-
- Spergel DJ, Kruth U, Shimshek DR, Sprengel R, Seeburg PH. Using reporter genes to label selected neuronal populations in transgenic mice for gene promoter, anatomical, and physiological studies. Prog Neurobiol. 2001;63(6):673–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
